
INTRODUCTION
During the seed development of fish, various morphological changes occur in body shape. These changes, referred to as metamorphosis, are species-specific, with external morphological changes occurring during the period of growth from hatching larvae to juvenile (McMenamin & Parichy, 2013). In fish species commonly cultured in Korea, for instance, the flounder (Paralichthys olivaceus) exhibits bilateral symmetry and a free-swimming lifestyle during the larval stage, similar to other fish species. However, as the larvae grow into juveniles and reach the benthic stage, the right eye migrates across the head to the left side, resulting in a body shape where one side faces the substrate (Kim & Kohno, 2009). In the case of the eel, the larvae, known as leptosephalus, are flat and transparent, and undergo a metamorphosis to become glass eels (Kim et al., 2014). For grouper species, during the larval stage, a distinctive morphological feature is the elongation of the second dorsal and pelvic fin spines. As they grow into juveniles, the fin lengths undergo changes, eventually resembling the proportions seen in adult fish (Kawabe & Kohno, 2009).
The metamorphosis process of fish seeds requires careful management during cultivation. With the recent push for diversification in aquaculture species both domestically and internationally, there has been increasing interest in developing aquaculture technologies for grouper species. Global grouper aquaculture production has seen significant growth, rising from 150,000 tons in 2010 to 227,000 tons in 2021, representing a 151% increase (FAO, 2024). This trend of increasing grouper production continues annually. Most grouper species are high-value fish, and the increasing market demand and profitability have led to a sustained increase in the demand for seed production and aquaculture. In the Asia-Pacific region, 60% of the grouper produced is traded in the Hong Kong market, with over 50% of that being re-exported to China. The red spotted grouper, in particular, is known to command the highest market price in Hong Kong (1,815.00 HKD/kg, Hong Kong Fish Market, 2024) and is recognized as a high-value aquaculture species, making it an attractive focus for aquaculture development.
In Korea, although seed production through artificial spawning has been successfully achieved to increase the aquaculture production of red spotted grouper (Lee et al., 2020), this species is known to be difficult for large-scale seed production due to high mortality rates during larval stages. Various studies on the aquaculture production of grouper species have been conducted, including research on the induction of maturation in red spotted grouper (Lee et al., 2020), feeding organisms (Boo et al., 2020), and egg development and larval morphological development (Dingerkus & Uhler, 1980; Hussain & Higuch, 1980; Kohno et al., 1993; Sawada et al., 1999; Kusaka et al., 2001; Leu et al., 2005; Song et al., 2005; Kawabe & Kohno, 2009; Park et al., 2014; Chen et al., 2021). Research on seed management and rearing related to the morphological development of red spotted grouper larvae is still insufficient.
This study aims to investigate morphological development characteristics of the fins during the larval stage to improve early survival in red spotted grouper seed production, and to obtain information on the management of appropriate rearing density management based on the behavioral characteristics of the larvae.
MATERIALS AND METHODS
The broodstock of red spotted grouper was reared in a recirculating aquaculture system (concrete rectangular tank 5.0×5.0×1.2 m, with an effective water volume of 25 tons), where the water temperature was maintained at 23.5±0.5°C, dissolved oxygen at 7.5±0.5 mg/L, and pH between 8.1–8.3. The broodstock were fed mackerel and other raw feed, as well as commercial feed, once in the morning and in the afternoon. The commercial feed was supplemented with multivitamins, B12, and omega-3, which were diluted in water and absorbed into the feed before being supplied.
The matured red spotted grouper broodstock were selected using cannulation. The selected fish were then intramuscularly injected with human chorionic gonadotropin (HCG) at a dosage of 500 IU per kilogram of body weight. Ovulation and sperm collection were induced 48 hours after the HCG injection by applying gentle pressure to the abdomen, leading to the collection of eggs and sperm. The fertilized eggs were produced using the wet method. Floating eggs were collected and stocked at a density of 200,000 individuals in a circular tank (Ø4.0×1.2 m, effective water volume of 12 tons).
The rearing conditions for the red spotted grouper seeds was maintained with dissolved oxygen levels 7.5-8.5 mg/L and pH range of 7.0-8.2. The water temperature was initially set at 21±0.1 °C and was gradually increased by 1°C per day after stocking the fertilized eggs, eventually stabilizing at 25.5±0.2°C. To enhance the nutrition of the rotifers supplied to the grouper seed rearing tanks, concentrated Chlorella (1.5×1010 cells/mL) was added to the tanks in the morning and afternoon. During the early stages of incubation, the rearing water was maintained in a static system, and after grading, the water flow rate was increased by transitioning to a flow-through system.
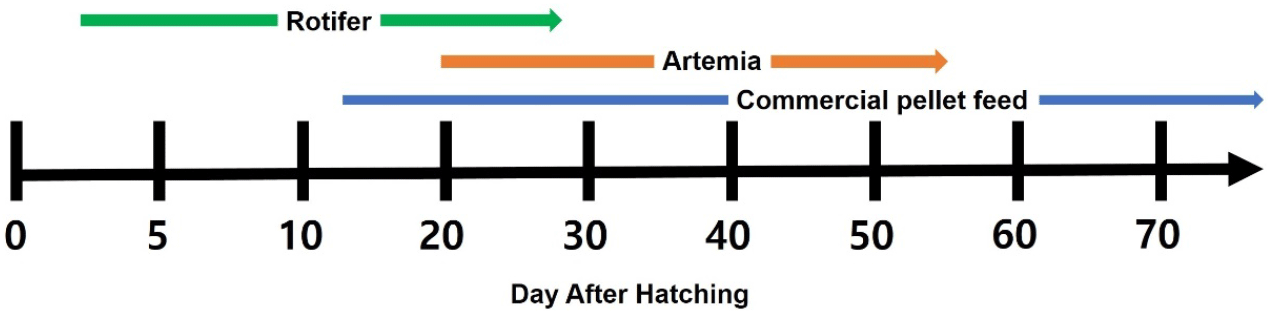
The feed for red spotted grouper seeds included rotifers, Artemia, commercial microparticulated feed. Rotifers were supplied from 3 DAH to 27 DAH. Newly hatched Artemia were provided from 20 DAH to 25 DAH, followed by nutritionally enriched Artemia from 26 DAH to 54 DAH. Commercial microparticulated feed was introduced in small amounts starting from 12 DAH, with the quantity gradually increasing from 20 DAH (Fig. 1).
To investigate the morphological changes in the fins of red spotted grouper larvae as they grow, fish were randomly sampled at 30 individuals each on days 1, 3, 5, 7, 9, 10, 11, 13, 15, 17, 19, 20, 25, 30, 39, 45, 51, and 70 DAH (day after hatching). The dorsal, pelvic, anal, and caudal fins of the larvae were observed using an optical microscope (OLYMPUS DP73, Tokyo, Japan) and a stereomicroscope (Zeiss Microscope Stemi SV 11 Apo Stereoscope, Oberkochen, Germany). The total length and the size of each fin were measured to an accuracy of 0.01 μm using Cellsens standard software and excope software. The length of the dorsal and pelvic fins was measured from the base or basal joint of the spine to the tip of the fin.
To investigate the skeletal development of the fins in red spotted grouper seeds, skeletal staining was conducted on experimental fish at 13, 20, 30, and 72 DAH, following the method outlined by Dingerkus & Uhler (1997). The experimental fish were fixed in 4% formalin for 24 hours. After fixation, the specimens were placed under running water for 24 hours to remove the formalin. Dehydration of the specimens was performed by immersing them sequentially in 50%, 70%, and 100% ethanol (EtOH) for 24 hours each.
The cartilage was stained by immersing the specimens in Alcian Blue solution for 24–48 hours. The Alcian Blue solution was prepared by adding 100 mg of Alcian Blue to 800 mL of 95% EtOH and 200 mL of glacial acetic acid. After staining, the specimens were neutralized in 100% EtOH for 24 hours and then rehydrated by immersing them in 70% and 50% EtOH at 2–3 hour intervals, with a final step in distilled water until the specimens sank.
The specimens were then placed in a 1% KOH solution for approximately one hour. For bone staining, the specimens were immersed in Alizarin Red S solution for 24 hours or until the bones appeared red or purple. After staining, the specimens were rinsed in a 1% KOH solution for 15–30 minutes. To remove melanin pigments, the specimens were bleached by immersing them in a bleaching solution for 1 to 1.5 hours. The bleaching solution was prepared by adding 3–4 drops of 3% hydrogen peroxide (H2O2) to 100 mL of 0.5% KOH solution.
RESULTS
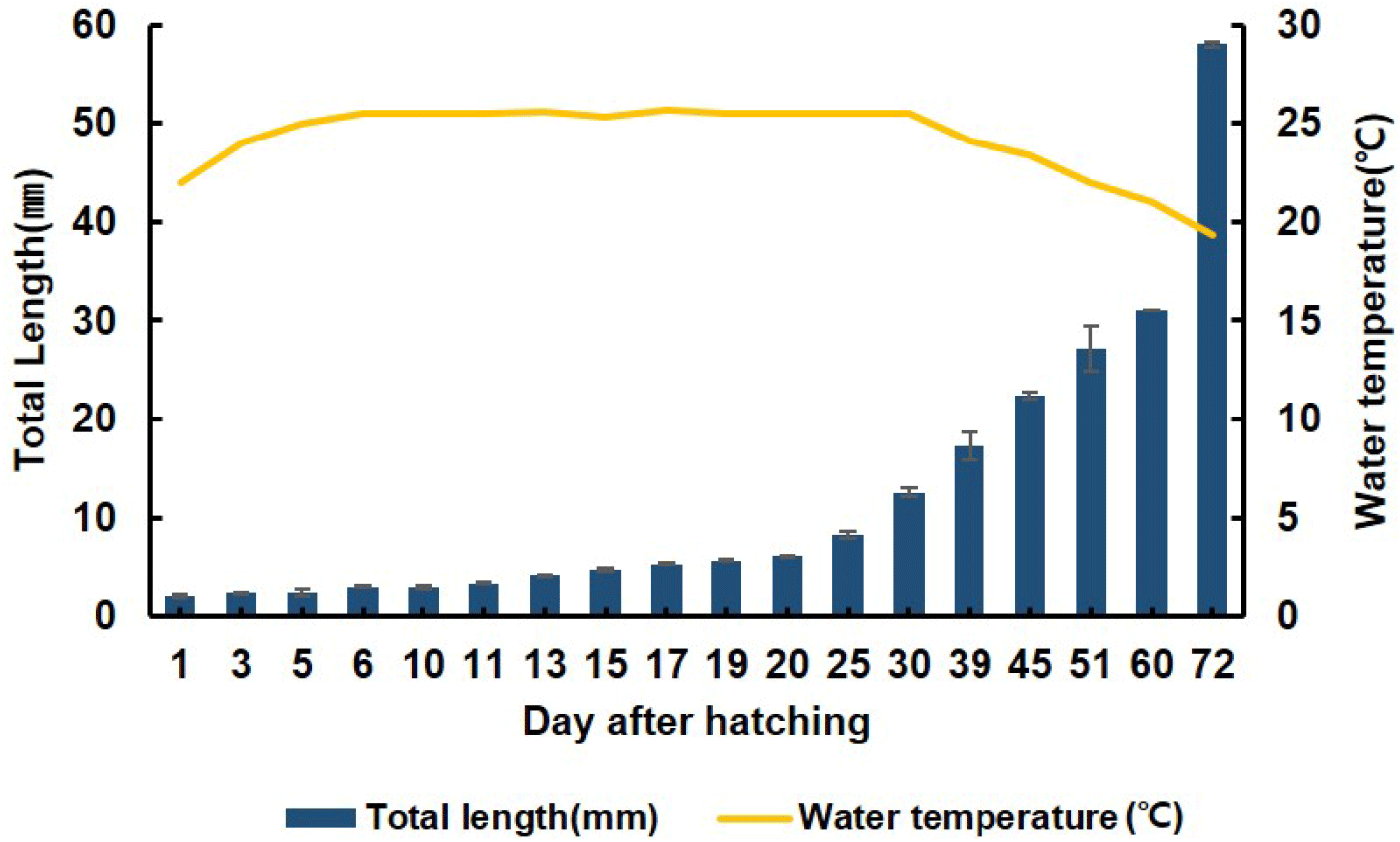
From 1 to 5 DAH, the average total length of the red spotted grouper larvae ranged from 2.1±0.04 to 2.41±0.03 mm. From 6 to 30 DAH, the average total length of the larvae increased to 2.62±0.05 to 12.6±0.37 mm. The rearing conditions for the seeds from 1 to day 30 DAH involved a stagnant system, with the water temperature maintained at 25.5±0.2°C. At 30 DAH, the rearing conditions were switched from a stagnant to a flow-through system. From 30 to 72 DAH, the average total length of the seeds increased to 12.6±0.4 to 56.0±0.2 mm, and water temperature was 19.3 °C (Fig. 2).
At 1 DAH of red spotted grouper larvae (average total length 2.1±0.04 mm), the mouth and anus were closed, yolk sac was oval-shaped, containing a single oil globule. The body was transparent, with a fin fold formed around the body (Fig. 3A). At 3 DAH of larvae (average total length 2.39±0.01 mm), their mouths and anuses open. The eyes had developed pigmentation, appearing silver, and the pectoral fins had formed into a fan shape (Fig. 3B). At 5 DAH, the larvae (average total length 2.41±0.03 mm) showed concentrated melanophores along the digestive tract and on the vertebrae above it, with melanophores distributed in a line along the ventral side of the tail (Fig. 3C). At 9 DAH, the larvae (average total length 2.74±0.1 mm) exhibited the emergence of pelvic fins near the ventral side, close to the digestive tract (Fig. 3D). At 10 DAH, the larvae (average total length 3.0±0.2 mm) showed the appearance of the second dorsal fin, measuring 0.2±0.03 mm, and began to exhibit schooling behavior in the rearing tank (Fig. 3E). At 11 DAH, the larvae (average total length 3.4±0.1 mm) had elongated second dorsal and pelvic fins, with a dot-like melanophore appearing at the rear end of the pelvic fins (Fig. 3F). At 13 DAH (average total length 4.2±0.1 mm), the first dorsal fin was observed, measuring 0.2±0.04 mm in length. Star¬shaped or dot-like melanophores were observed at the rear ends of the second dorsal and pelvic fins (Fig. 3G). At 17 DAH, the larvae (average total length 5.3±0.1 mm) exhibited a 45° curvature of the caudal region, with the third dorsal fin emerging and measuring 0.22 mm in length (Fig. 3H). At 17 to 19 DAH, the length of the pelvic fins reached their maximum growth relative to the total length, ranging from 1.62±0.08 to 1.79±0.05 mm (Fig. 3I). At 19 to 20 DAH, the second dorsal fin also reached its maximum growth relative to the total length, measuring 1.95±0.11 to 2.11±0.04 mm. At 20 DAH (average total length 6.1±0.1 mm), the fourth dorsal fin was observed (Fig. 3J). At 30 DAH (total length 12.6±0.4 mm), the larvae reached the complete count of fin rays in each fin: 11 spines and 14-15 soft rays in the dorsal fin, 2 spines and 10 soft rays in the pelvic fin, 3 spines and 8 soft rays in the anal fin, and 13 caudal fin rays (Fig. 3K). At 39 DAH, the juvenile of red spotted grouper (average total length 20.28±3.1 mm) had developed a body shape similar to that of adult fish and displayed active schooling behavior, swimming in circles in the rearing tank (Fig. 3L). At 51 DAH (average total length 30.8±2.3 mm), all fins had turned yellow. The body color had developed into a brown base with small, round spots distributed across the entire body (Figs. 3M, 4, and 5).

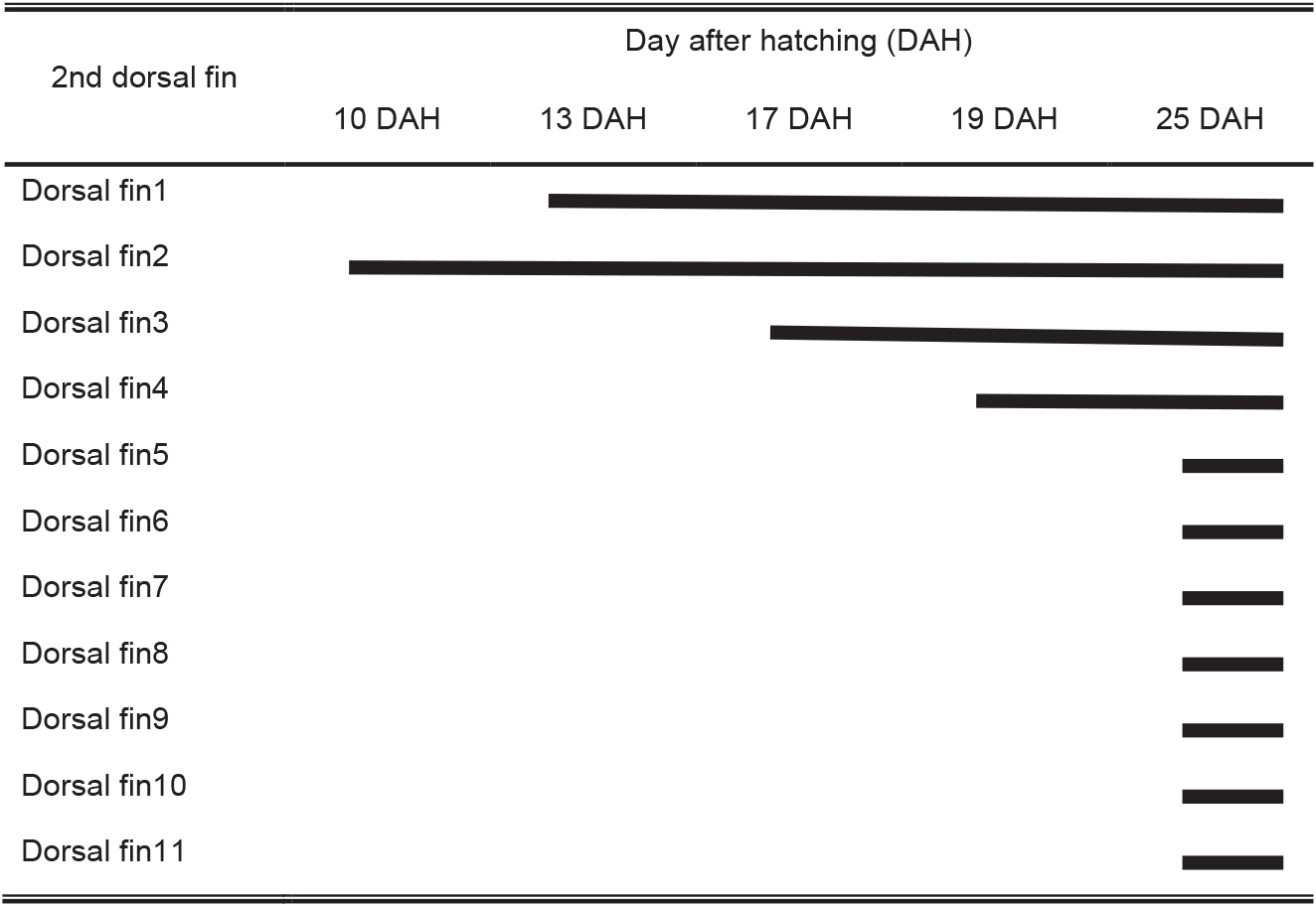
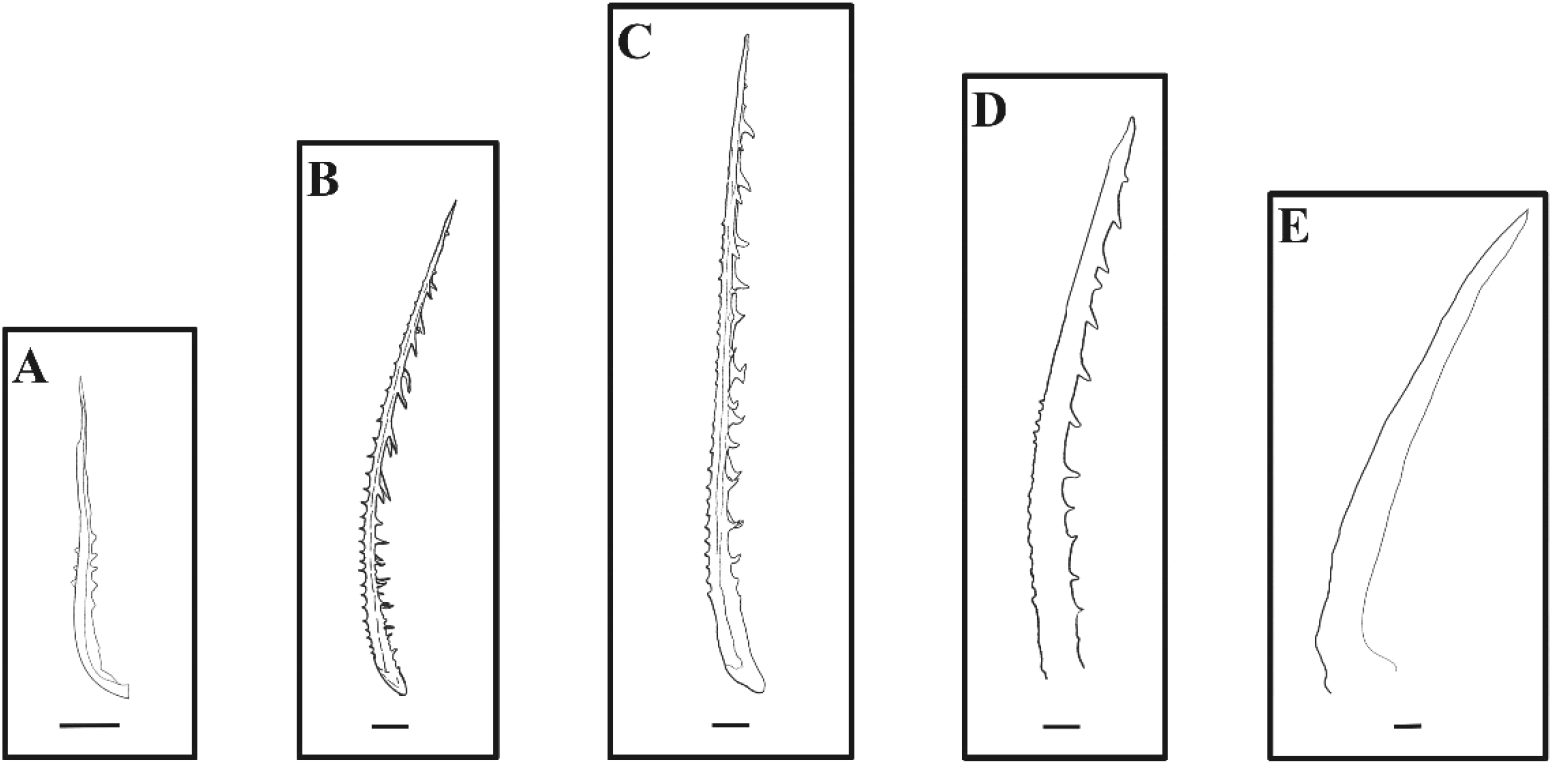
The number, length, and external morphology of the barb and spinule in the second dorsal fin of red spotted grouper seeds were investigated. The second dorsal fin appeared at 10 DAH in larvae (3.0±0.2 mm TL). This fin consisted of one apex ridge and two posterolateral wings. At 13 DAH (4.2±0.1 mm TL), barbs and spinules were first observed in the second dorsal fin. The barbs and spinules formed along the anterior and posterior ridges (apex ridge and posterolateral wings) of the second dorsal fin, with an average of 7.8±2.7 barbs and 2.4±0.9 spinules, and their lengths were 9.5±2.3 μm and 2.5±1.4 μm, respectively. At 20 DAH (6.1±0.1 mm TL), the number and length of the barbs and spinules on the second dorsal fin ridges had increased, with 25±2.7 barbs and 25.8±2.4 spinules observed, measuring 50.4±6.0 μm and 27.1±1.7 μm, respectively. At 30 DAH (12.6±0.4 mm TL), the number of barbs and spinules was 27±1.4 and 23.3±2.8, with lengths of 67.5±3.7 μm and 16.3±2.8 μm, respectively. However, at 39 DAH (20.3±3.1 mm TL), the number of barbs and spinules had decreased to 22.7±3.7 and 13±4.6, respectively. The length of the barbs decreased to 45.8±3.7 μm, while the length of the spinules increased to 34.5±11.1 μm. At 72 DAH (56.0±0.2 mm TL), these barbs and spinules were no longer observed on the second dorsal fin (Fig. 6, Table 1).
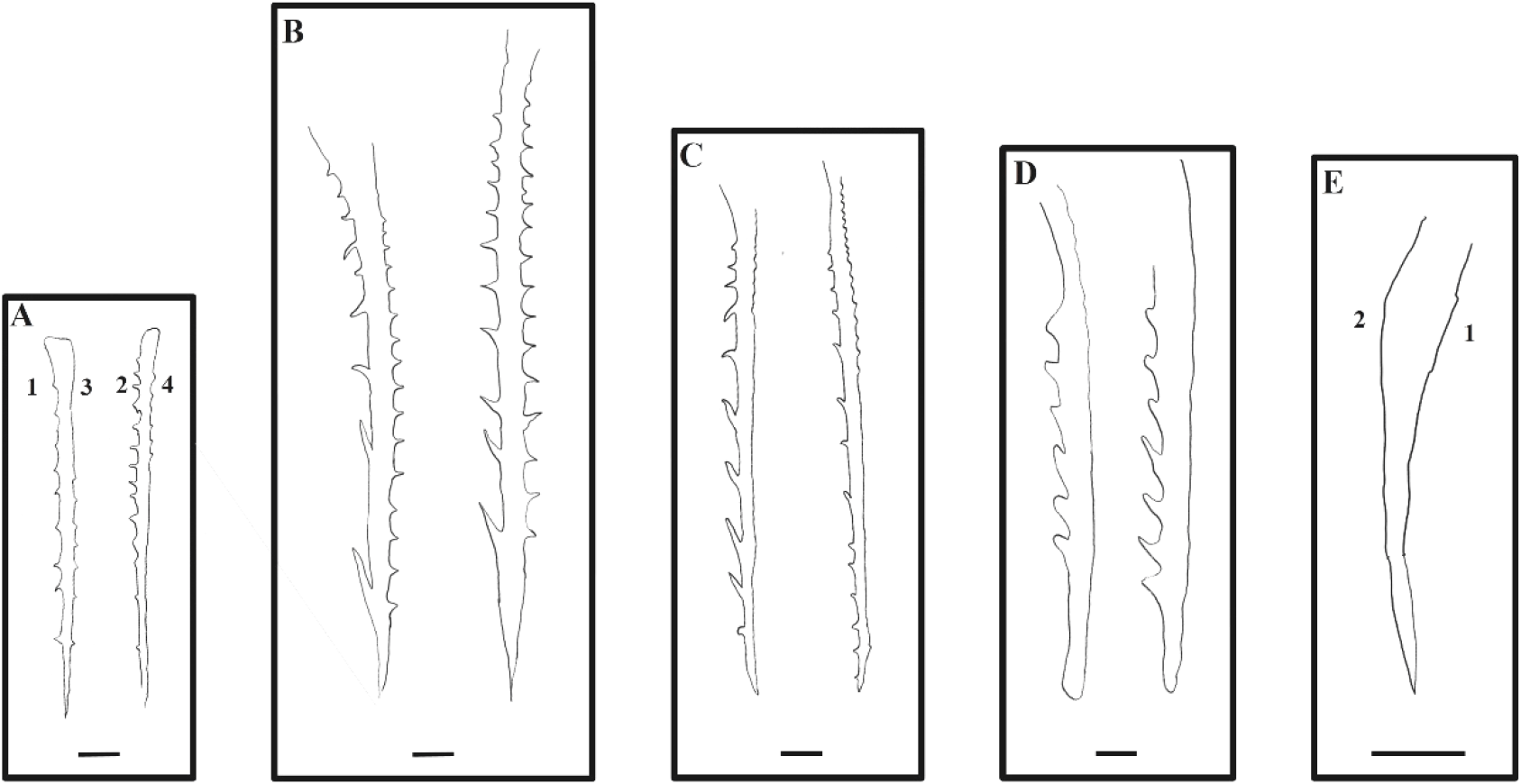
The number, length, and external morphology of the barb and spinule in the pelvic fins of red spotted grouper seeds were examined. At 9 DAH (3.0±0.2 mm TL), the pelvic fins first appeared, with four distinct ridges developing (Dorsomedial ridge, Ventromedial ridge, Ventrolateral ridge, Dorsolateral ridge). At 13 DAH (4.2±0.1 mm TL), barbs and spinules were first observed in the pelvic fins. The number of barbs formed on ridge 1 and ridge 2 of the pelvic fins were 3.2±1.6 and 6.8±2.0, respectively, with an average length of 10.8±1.7 μm for each. Spinules were observed on ridge 3 of the pelvic fins, with an average of 1.2±1.1 spinules measuring 1.0±0.9 μm in length. At 20 DAH (6.08±0.07 mm TL), the number of barbs on ridges 1 and 2 of the pelvic fins had increased to 12.3±2.3 and 14.3±0.9, respectively, with average lengths of 44.8±8.7 μm and 39.6±4.5 μm. Spinules were observed on ridges 3 and 4, with 19.3±2.5 and 21±1 spinules, and average lengths of 16.5±2.7 μm and 15±1 μm, respectively, showing an increase in both number and length compared to 13 DAH. At 30 DAH (12.6±0.4 mm TL), the number of barbs on ridges 1 and 2 of the pelvic fins was 9.7±0.7 and 10.7±1.2, with average lengths of 56.7±10.5 μm and 34.79±0.95 μm, respectively. The number of spinules on ridges 3 and 4 was 9.5±5.5 and 11.04±6.38, with average lengths of 13.2±1.3 μm and 3.2±2.6 μm, respectively. At 39 DAH (20.3±3.1 mm TL), the number of barbs on ridges 1 and 2 was reduced to 3.3±2.7 and 11.3±1.5, with average lengths of 24.2±19.7 μm and 63.3±17.3 μm, respectively. Spinules were only observed on ridge 3, with an average number of 9.5±5.5 and a length of 23.6±2.3 μm. At 72 DAH, these barbs and spinules were no longer observed on the pelvic fins (Fig. 7, Table 2).
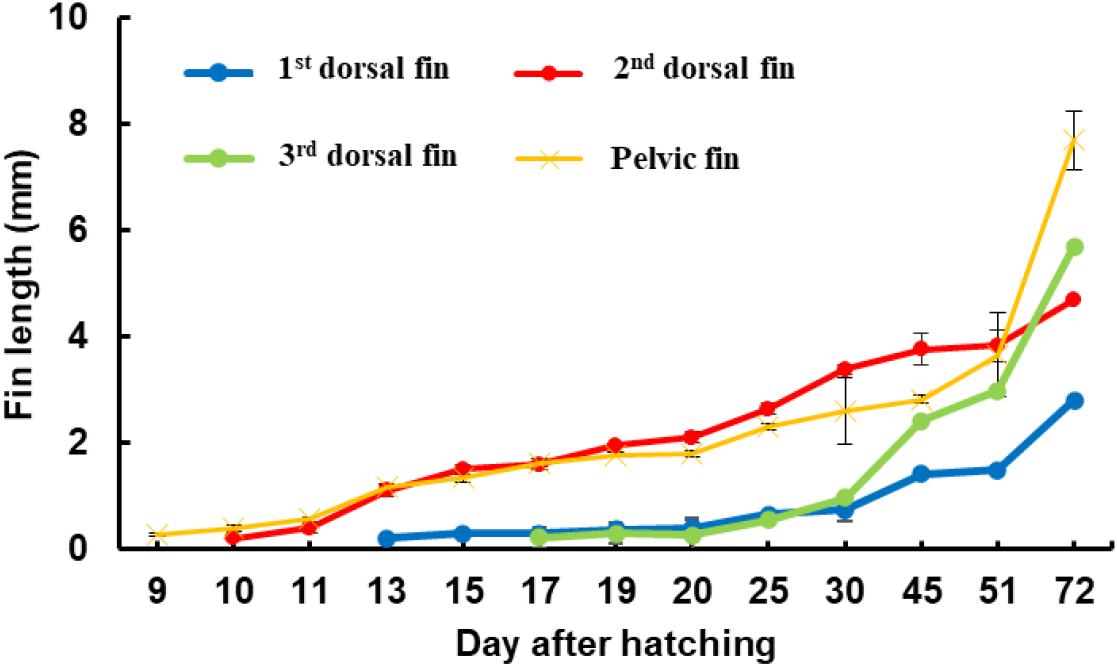
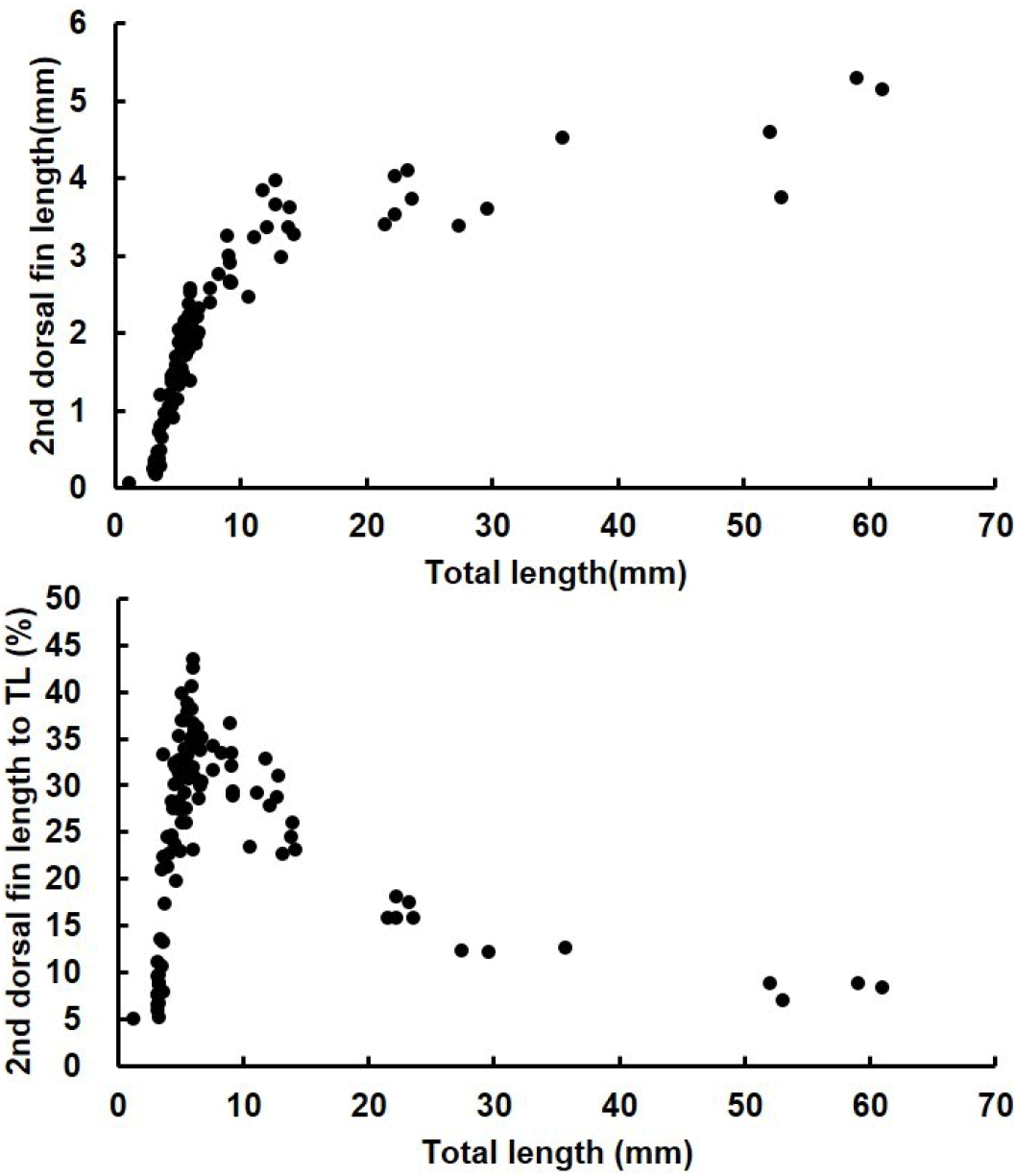
The length of the second dorsal fin relative to the total length was investigated in red spotted grouper ranging from 1.16 to 61.0 mm TL. At 10 DAH, when the second dorsal fin first appeared, the average length of the dorsal fin was 0.2±0.03 mm. The length of the second dorsal fin grew rapidly, reaching an average of 3.38±0.1 mm at 30 DAH (12.6±0.4 mmm TL). However, from 30 DAH to 72 DAH, as the seeds grew from an average TL of 12.6±0.4 mm to 56±0.2 mm, the average length of the second dorsal fin increased more gradually, ranging from 3.38±0.1 mm to 4.7±0.3 mm. After 30 DAH, the rate of growth in the second dorsal fin length slowed compared to earlier stages but continued to increase with overall growth. At 20 DAH (6.1±0.1 mm TL), the ratio of the second dorsal fin length to total length reached its peak at 34%. As the seeds continued to grow, this ratio decreased, with the second dorsal fin length relative to total length being 16% at 45 DAH (average TL 22.5±0.3 mm) and 9% at 72 DAH (average TL 56.0±0.2 mm), indicating a decreasing trend as the seeds grew (Fig. 8).
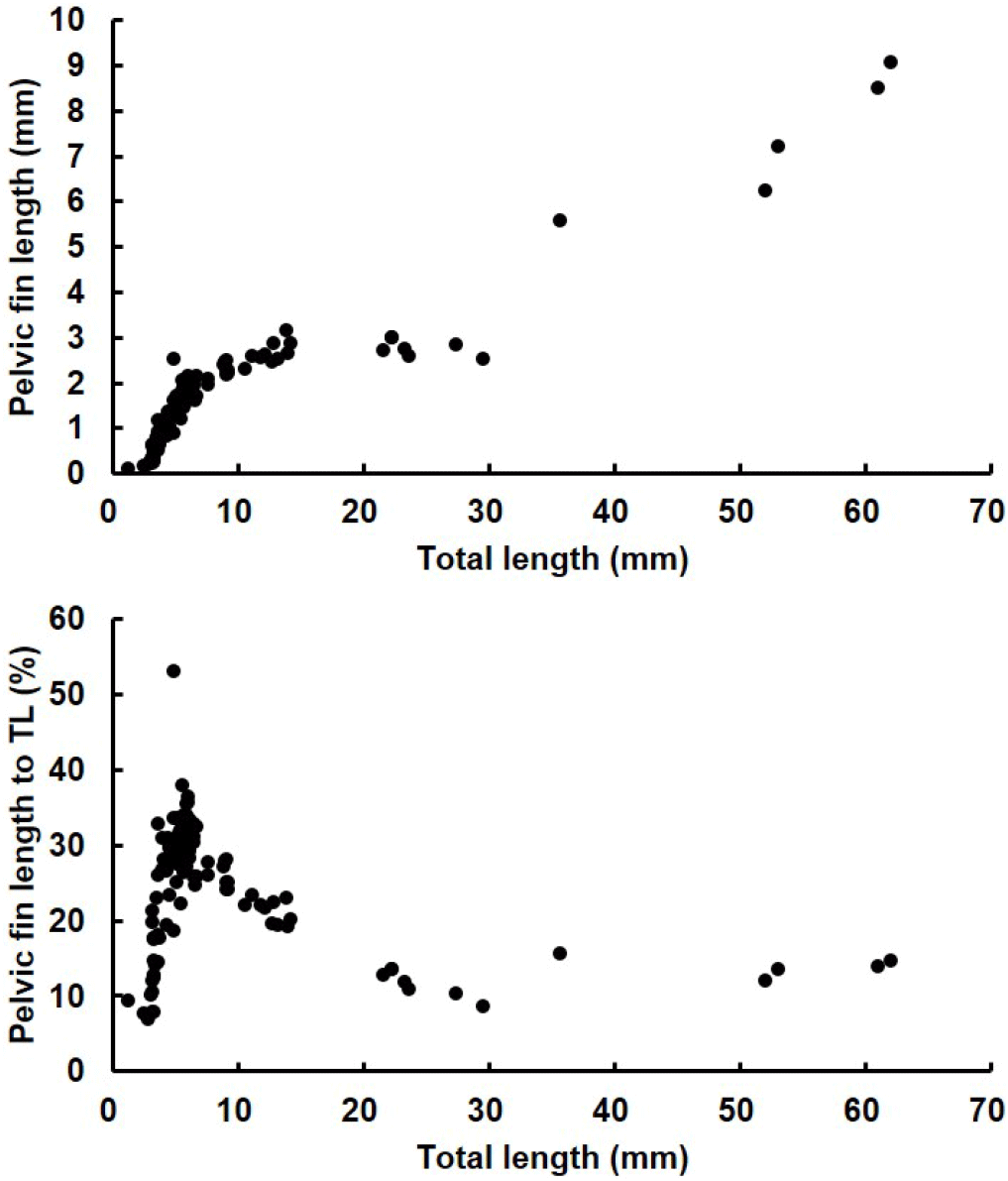
The length of the pelvic fin spines relative to total length was investigated in red spotted grouper ranging from 1.16 mm to 62.0 mm TL. At 9 DAH, when the pelvic fins first appeared, the average length of the pelvic fins was 0.26±0.03 mm. The pelvic fins grew rapidly, reaching an average length of 1.80±0.05 mm at 20 DAH (6.1±0.1 mm TL). From 20 DAH to 51 DAH, as the seeds grew from an average TL of 6.1±0.1 mm to 30.84±2.02 mm, the pelvic fin length ranged from 1.8±0.1 mm to 3.7±0.8 mm.
At 19 DAH, the average percentage of pelvic fin length relative to total length reached its peak at 31% in larvae with an average TL of 5.7±0.1 mm. However, this percentage decreased as the larvae continued to grow, with the average percentage of pelvic fin length relative to total length being 21% at 30 DAH (12.6±0.4 mm TL) and 12% at 51 DAH (30.8±2.3 mm TL), indicating a declining trend as the seeds grew (Fig. 9).
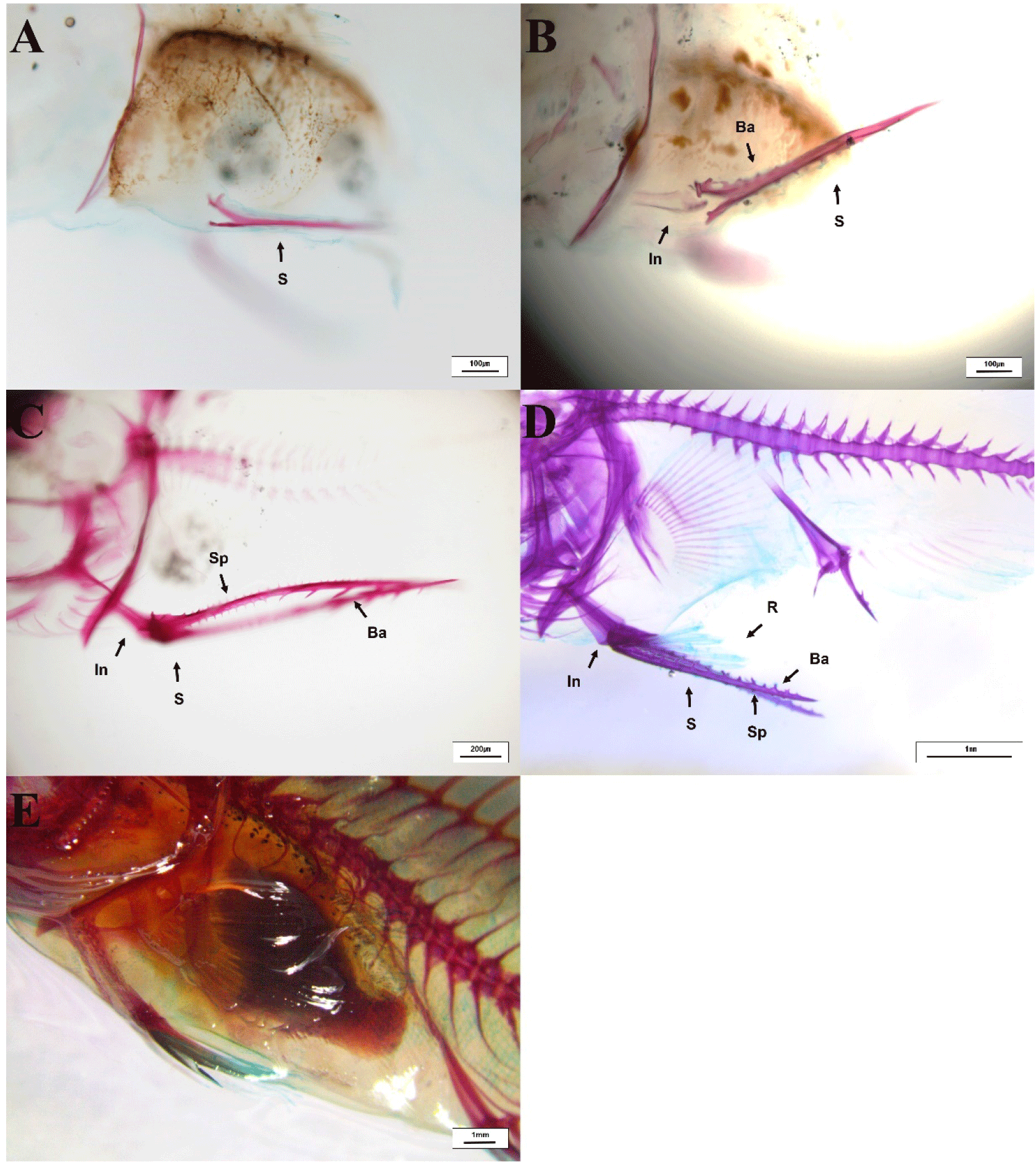
At 13 DAH (4.2±0.1 mm TL), the second dorsal fin began to ossify, and the first dorsal fin also appeared and started ossification (Fig. 10A). At 20 DAH (6.08±0.07 mm TL), the proximal radials of both the first and second dorsal fins were ossified, and the third dorsal fin had appeared and begun ossification. All fin spines were fully ossified at this stage (Fig. 10B). At 33 DAH (12.6±0.9 mm TL), nine dorsal fins were observed to be ossified, and the ossification of 11 proximal radials and the fin rays was completed. Additionally, the distal radials of the dorsal fin rays were also fully ossified (Fig. 10C and 10D). At 72 DAH (56±0.2 mm TL), the fins of the red spotted grouper seeds were entirely ossified (Fig. 10E and 10F).
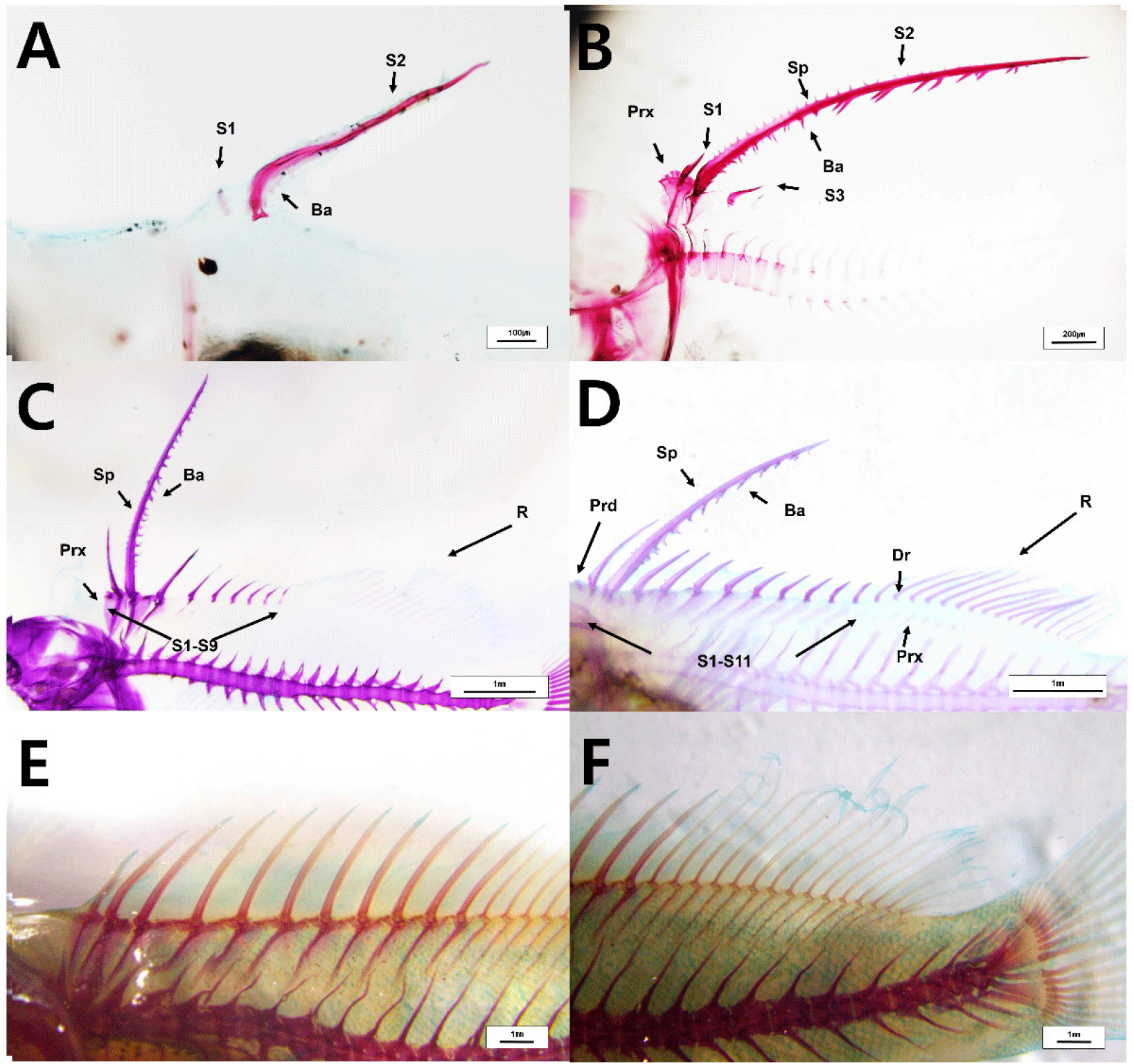
At 13 DAH (4.2±0.1 mm TL), the pelvic fin spine and the rod-shaped innominatum were in the process of ossification (Fig. 11A and 11B). At 20 DAH (6.1±0.1 mm TL), the pelvic fin spines and rays were fully ossified (Fig. 11C and 11D). At 30 DAH (12.6±0.4 mm TL), the ossification of the innominatum in the pelvic fin was completed (Fig. 11E).
DISCUSSION
Certain fish species undergo various morphological changes during seed development. In the case of the red spotted grouper, a distinctive feature of its seed development is the unique elongation of the second dorsal fin and pelvic fin spines during the metamorphosis process as it grows into a juvenile. These morphological changes are critical factors in determining appropriate rearing densities during the seed production process. This study investigated the morphological changes in the fins of red spotted grouper seeds throughout their growth to provide information on optimal rearing density management.
In red spotted grouper larvae, two pelvic fin spines and one second dorsal fin spine appeared at 10 DAH (3.0±0.2 mm TL), and their length began to increase rapidly. At 13 DAH (4.2±0.1 mm TL), barbs and spinules began to emerge along the anterior and posterior ridges (Apex ridge and Posterolateral wings) of the dorsal fin. This pattern of spine development in red spotted grouper larvae is similar to that observed in other grouper species. For instance, in the Malabar grouper (Epinephelus malabaricus), the second dorsal fin and pelvic fin spines began to elongate at 9 DAH (3.78–4.23 mm TL), and saw-like barbs and spinules appeared on the fins at 14 DAH (5.23–5.61 mm TL) (Leu et al., 2005). Similarly, in the blacktip grouper (Epinephelus fasciatus), the spines of the second dorsal and pelvic fins emerged and elongated at 10 DAH (3.8 mm TL), and barbs and spinules were observed on these fins at 13 DAH (4.3 mm TL) (Kawabe & Kohno, 2009). Additionally, in the orange-spotted grouper (Epinephelus tauvina), the second dorsal and pelvic fins appeared and began to elongate at 10 DAH (3.8 mm TL), with barbs and spinules becoming visible on these fins at 13 DAH (4.3 mm TL) (Dingerkus & Uhler, 1980).
The measurements of the ratio of the second dorsal fin and pelvic fin lengths relative to the total length in red spotted grouper larvae showed that, at 19–20 DAH (average total length was 5.7±0.1 mm to 6.1±0.1 mm), the ratio of the second dorsal fin length averaged 33%–34%, while the ratio of the pelvic fin spine length averaged 30%–31%, both representing the highest values observed. Notably, at 19 DAH, the ratio of the second dorsal fin length relative to total length reached 43.53%, and at 25 DAH, the ratio of the pelvic fin length relative to total length reached its peak at 53.01%. Kusaka et al. (2001) reported that when red spotted grouper larvae reached a total length of 3.3 mm, the ratios of the second dorsal fin and pelvic fin lengths to total length were 45% and 44%, respectively. In the sevenband grouper (Epinephelus septemfasciatus), the highest relative ratios of the dorsal and pelvic fins were observed at around 75% and 54%, respectively, when the total length was approximately 8.1 mm (Kitajima et al., 1991). For E. malabaricus, the ratios were 53% and 49%, and for Epinephelus tauvina, they were 50% and 43% (Table 3). During the early development of red spotted grouper larvae, the second dorsal and pelvic fins, which elongate significantly relative to total length, represent species-specific traits of groupers. Rather than the fins shrinking during the metamorphosis process, they appear proportionally smaller as the juveniles grow in total length. Additionally, the ratios of the second dorsal fin and pelvic fin lengths to total length vary among grouper species, reflecting species-specific differences.
| Species | Second-dorsal fin | Pelvic fin |
Water temperature (°C) |
References | ||||
|---|---|---|---|---|---|---|---|---|
|
To TL (%) |
DAH |
TL (mm) |
To TL (%) |
DAH |
TL (mm) |
|||
| Epinephelus akaara | 43.53 | 19 | 5.95 | 53.01 | 25 | 4.81 | 25.3°C-25.7°C | This study |
| Epinephelus akaara | 45 | - | 3.3 | 44 | - | 3.3 | 24.7°C-27.3°C | Kusaka et al. (2001) |
| Epinephelus malabaricus | 52.68 | 20 | 7.39 | 48.62 | 20 | 7.39 | 26.1°C-26.8°C | Leu et al. (2005) |
| Epinephelus fasciatus | 51 | - | - | 41 | - | - | 22.5°C-27.0°C | Kawabe & Kohno (2009) |
| Epinephelus bruneus | 40 | - | 8 | 35 | - | 8 | - | Sawada et al. (1999) |
| Epinephelus tauvina | 50 | - | 8 | 43 | - | 6 | - | Hussain & Higuch (1980) |
| Epinephelus septemfasciatus | 75 | 33 | 8.1 | 54 | 33 | 8.1 | 22.0°C-28.0°C | Kitajima et al. (1991) |
In this study, barbs and spinules were observed to form on the second dorsal fin and pelvic fin spines of red spotted grouper larvae. At 13–20 DAH, the length and number of these barbs and spinules increased rapidly. However, at 39 DAH, the number of barbs and spinules on the fins showed a decreasing trend, and they were no longer observed at 72 DAH. A similar pattern was reported in the blacktip grouper (E. fasciatus), where sharp barbs and spinules formed on the anterior and posterior parts of the second dorsal fin when the fish reached a total length of 5.18 mm. The barbs and spinules gradually became smaller, and by the time the fish reached 40.2 mm in total length, the barbs and spinules had degenerated (Kawabe et al., 2009).
The appearance of barbs and spinules on the second dorsal fin and pelvic fin spines during the larval stage of grouper species is likely a crucial factor in determining the appropriate rearing density during the seed production process. The red spotted grouper larvae tend to exhibit schooling behavior in the morning, and during this time, there is a risk of mortality due to entanglement of the barbs and spinules on the second dorsal and pelvic fin spines. Therefore, careful attention should be given to managing the behavior patterns and rearing density of red spotted grouper larvae from the time these barbs and spinules appear until the metamorphosis is complete and the spines degenerate.