
INTRODUCTION
Nucleobindin protein, which was first identified in human and mouse cell lines, has two isotypes: nucleobindin 1 (NUCB1) and nucleobindin 2 (NUCB2) (Barnikol-Watanabe et al., 1994; Miura et al., 1992). However, NUCB2 only functions physiologically in humans and rodents (Miura et al., 1992). Nesfatin-1, nesfatin-2, and nesfatin-3 are generated through post-translational processing of NUCB2 via the enzyme pro-hormone convertase-1/3 (Gonzalez et al., 2010; Oh-I et al., 2006). A physiological activity has only been demonstrated for nesfatin-1 (Gonzalez et al., 2010). Nesfatin-1 is expressed in various hypothalamic nuclei for appetite control such as the arcuate nucleus (ARC), paraventricular nucleus (PVN), supraoptic nucleus (SO), lateral hypothalamic area (LHA), and zona incerta in rats (Brailoiu et al., 2007; Fort et al., 2008; García-Galiano et al., 2010; Goebel et al., 2009; Kohno et al., 2008). Nesfatin-1 also exists in cerebrospinal fluid and the brain stem, including the locus coeruleus, rostral raphe pallidus, Edinger-Westphal nucleus, and ventrolateral medulla (Goebel et al., 2009; Stengel et al., 2010). Intracerebroventricular infusion of nesfatin-1 decreases food intake and inhibits feeding behavior (Atsuchi et al., 2010; Shimizu et al., 2009), whereas infusion of a nesfatin-1-neutralizing antibody stimulates appetite (Oh-I et al., 2006).
Recent studies identified nesfatin-1 immunoreactivity in rat gastric organs (Gonzalez et al., 2009; Stengel et al., 2010). Nesfatin-1 is highly expressed in gastric endocrine cells and duodenal Brunner’s glands of rodents (Stengel et al., 2009). X/A-like cells secrete ghrelin, which stimulates appetite, in the gastric mucosal layer, and these cells also express nesfatin-1 (Stengel et al., 2010). These results raise the possibility that nesfatin-1/NUCB2 gene expression is regulated by nutritional status, suggesting a regulatory role of peripheral nesfatin-1 in energy homeostasis. Nesfatin-1 immunoreactive cells co-localize with insulin-containing pancreatic β-cells in mice and rats (Gonzalez et al., 2009), suggesting a role for nesfatin-1 in pancreatic islet and glucose homeostasis (Foo et al., 2010).
Furthermore, nesfatin-1 is detected in the fat of humans and rodents and is expressed more in subcutaneous fat than in visceral fat (Ramanjaneya et al., 2010). In experiments with fat cell lines, nesfatin-1 expression increases exponenttially during the differentiation of fat progenitor cells into mature fat cells (Ramanjaneya et al., 2010). The fact that the nesfatin-1 concentration is high in the blood of people with a high body mass index demonstrates that nesfatin-1 secreted from fat cells may regulate food intake independently of leptin (Ramanjaneya et al., 2010; Shimizu et al., 2009).
Several recent reports demonstrated that nesfatin-1 protein is expressed in the reproductive system. Nesfatin-1 localizes in interstitial cells, including Leydig cells, in rat and mouse testis (García-Galiano et al., 2010). In addition, nesfatin-1 binding sites were detected on theca cells and interstitial cells near the tunica albuginea of mouse ovary, and on boundary cells in the tunica albuginea of mouse testis (Kim et al., 2010; Kim et al., 2011). Many studies have reported the expression of nesfatin-1/NUCB2 mRNA and protein in various tissues. Therefore, the purpose of this study was to assess the expression levels and distributions of nesfatin-1/NUCB2 in various mouse tissues.
MATERIALS AND METHODS
Six-week-old ICR male and female mice were purchased from Samtako Bio Korea (South Korea) and housed in groups of five per cage under controlled illumination (12:12 h light/dark cycle, lights on/off: 6 h/18 h) and temperature (22 ± 2°C). Animals were fed a standard rodent diet and tap water ad libitum. Mice were euthanized by CO2 anesthesia followed by cervical dislocation. Cerebrum, hypothalamus, pituitary, stomach, heart, liver, intestine, spleen, lung, thymus, ovary, uterus, testis, epididymis, fat and muscle were quickly removed and stored. Animal care and experimental procedures were approved by the Institutional Animal care and the use committee at the Seoul Women’s University in accordance with guidelines established by the Korea Food and Drug Administration.
Total RNA was isolated by using the RNA isoplus (TaKaRa Bio, Shiga, Japan) according to manufacturer’s instruction. The final pellet was air dried and dissolved into RNase-free DEPC solution (TaKaRa Bio, Shiga, Japan). The RNA concentration was measured with the Nano-drop (Thermo Fisher Scientific Inc., Waltham, MA). First strand cDNA synthesis was performed in RNase-free DEPC solution containing 2 μg total RNA and 10 pmol oligo dT at 70°C for 5min, followed by double-strand synthesis in 5X RT buffer (Invitrogen, Carlsbad, CA) with 8 mM dNTP (BIO BASIC INC., Ontario, Canada), 200 unit/μL RTase (Invitrogen, Carlsbad, CA) at 37°C for 60 min and at 72°C for 15 min.
Conventional PCR was performed in buffer solution containing 3 μL of template cDNA, 5 unit/μL of Taq polymerase (BIONICS, Korea), 0.25 mM dNTPs (BIO BASIC INC., Ontario, Canada) and 10 pmol of each primer. Primers were designed for NUCB2 and β-actin on the basis of the mouse cDNA sequences. The following primer pairs were used: NUCB2 forward 5'-TTTGAACACCTGAACCACCA- 3'; reverse 5'-TGGTCTTCGTGCTTCCTCTT-3' and β- actin forward 5'-CTCTTTGATGTCACGCACGATTTC-3'; reverse 5'-ATCGTGGGC CGCTCTAGGCACC-3' primers (BIONICS, Korea). The optimum temperature cycling protocol was used as 95°C for 15s, 60°C for 30s and 72°C for 30min, using the GenePro thermal cycler (Bioer, China). The reaction products were run on a 2% agarose gel and visualized with ethidium bromide to check the length of the amplified cDNA.
The tissues were homogenized in ice-cold lysis buffer 50 mM Tris-base (pH 7.4), 150 mM NaCl, 10 mM EDTA, 0.1% Tween-20, and protease inhibitors (0.1 mM phenylmethylsulfonylfluoride, 5 μg/mL aprotinin, and 5 μg/mL leupeptin). The homogenates were centrifuged at 12,000 × g for 30 min at 4°C. The protein concentration in the supernatant was determined by the DC protein assay kit (Bio-Rad Laboratories, Inc., Hercules, CA). Equal amounts of proteins (20 μg) were resolved by 12% SDS-PAGE and transferred to PVDF membranes (Amersham; GE Healthcare, Buckinghamshire, England). The membranes were then incubated in blocking solution with 3% Casein Blocking Solution (KOMABIOTECH, Korea) in Tris-buffered saline (TBS) containing 10 mM Tris (pH 7.6), 150 mM NaCl, and 0.1% Tween-20 for 1 h at room temperature. Then, the membranes were incubated with rabbit anti-rat nesfatin-1 polyclonal antibody (Phoenix Pharmaceuticals, INC., Burlingame, CA) and anti-mouse β-actin antibody (Santa Cruz Biotechnology, Inc., Paso Robles, CA) overnight at 4°C. The membrane was washed three times with washing buffer and incubated with goat anti-rabbit IgG-HRP (Santa Cruz Biotechnology, Inc., Paso Robles, CA) and donkey antimouse IgG-HRP (Santa Cruz Biotechnology, Inc., Paso Robles, CA) at room temperature for 1 h, respectively. After washing three times, the membrane was detected by ECL Plus Western Blotting Detection Reagents (Amersham; GE Healthcare, Buckinghamshire, England). The relative protein levels were analyzed by Scion Image (National Institutes of Health, Bethesda, MD).
Real-time PCR was performed in a total volume of 20 μL buffer solution containing 2 μL of template cDNA, 10 μL of SYBR Green (Roche, Manheim, Germany), and 10 pmol of each primer. Primer pairs were as follows: NUCB2 forward 5'-AAAACCTTGGCCTGTCTGAA-3'; reverse 5'- CATCGATAGGAACAGCTTCCA-3' and GAPDH forward 5'-TTGATGGCAACAATCTCCAC-3'; reverse 5'-CGTCC CGTAGACAAAATGGT-3' (BIONICS, Korea). The optimum temperature cycling protocol was determined to be 95°C for 5 min followed by 45 reaction cycles at 95°C for 10 s, 60°C for 10 s and 72°C for 10 s using the LightCycler® 480 Real-time PCR System (Roche, Manheim, Germany).
The hypothalamus, uterus, ovary and testis were removed and fixed in 4% paraformaldehyde for 2 h. The tissues were rinsed in ethanol series to remove fixative residues, embedded in paraffin block. The tissues blocks were cut 10 μm sections using a microtome, deparaffinized, and rehydrated with graded xylene-alcohol series, and then washed with PBS before immunostaining. The sections were blocked with peroxidase blocking reagent (Dako, Glostrup, Denmark) for 10 min and 2.5% horse serum for 20 min in order to prevent nonspecific binding. After rinsed with washing buffer, the sections were incubated with rabbit anti-rat nesfatin-1 polyclonal antibody (Phoenix Pharmaceuticals, INC., Burlingame, CA) at 4°C for overnight. Sections were rinsed and incubated with biotinylated goat anti rabbit IgG (Vector Laboratories, Burlingame, CA) followed by horseradish peroxidase-conjugated streptavidin (Vectastain Elite ABC reagent; Vector Laboratories, Burlingame, CA) for 30 min, respectively, at room temperature. Immunoreactivity was visualized using 3,3’-diaminobenzidine tetrahydrochloride (Dako, Glostrup, Denmark) and counterstained with Mayer’s hematoxylin (Sigma, St. Louis, MO). The sections were mounted on the slides with permounting media (Thermo Fisher Scientific Inc., Waltham, MA) and observed under light microscope (Axioskop2, Carl Zeiss, Germany).
RESULTS
We investigated NUCB2 mRNA expression by conventional PCR to determine whether the NUCB2 gene is expressed in the cerebrum, hypothalamus, pituitary, stomach, heart, liver, intestine, spleen, lung, thymus, ovary, uterus, testis, epididymis, fat, and muscle of mice. In female mice, NUCB2 mRNA was detected in the cerebrum, hypothalamus, pituitary, stomach, heart, liver, intestine, spleen, lung, thymus, fat, muscle, ovary, and uterus (Fig. 1A). Similarly, NUCB2 mRNA was expressed in all tested tissues of male mice, including the testis and epididymis (Fig. 2A).
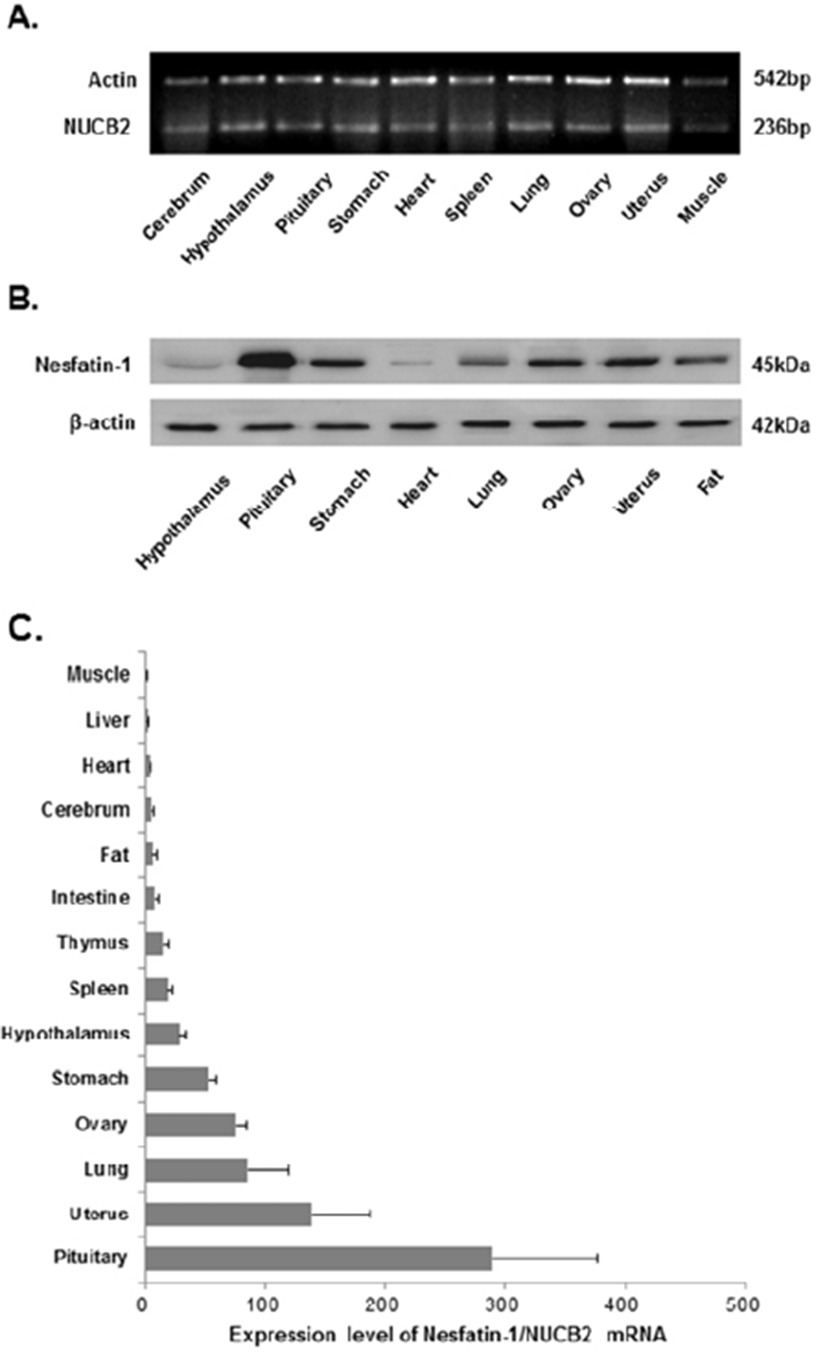

Western blotting detected nesfatin-1 protein expression in the hypothalamus, pituitary, stomach, heart, lung, ovary, uterus, testis, epididymis, and fat. In both female and male mice, nesfatin-1 protein was predominantly detected in the pituitary and at lower levels in the hypothalamus, spleen, thymus, heart, liver, and muscle. Expression was much higher in reproductive organs, such as the testis, epididymis, ovary, and uterus, than in the hypothalamus (Fig. 1B, 2B).
The relative expression levels of NUCB2 mRNA in the cerebrum, hypothalamus, pituitary, stomach, heart, liver, lung, intestine, spleen, ovary, uterus, testis, epididymis, muscle, and fat were analyzed by real-time PCR. The levels of NUCB2 mRNA expression in various tissues of female and male mice were similar. A large amount of NUCB2 mRNA was detected in the pituitary, in comparison with other organs. NUCB2 mRNA was also highly expressed in the stomach, lung, and reproductive organs (Fig. 1C, 2C).
The sites of nesfatin-1 protein expression were investigated by immunohistochemical staining in sections of the hypothalamus, uterus, ovary, and testis. Nesfatin-1 protein was localized in the PVN of the hypothalamus as a positive control (Fig. 3A, B). In the uterus, nesfatin-1 protein expression was detected in epithelial cells around the uterine glands and endometrium (Fig. 3D, E). In the ovary, nesfatin-1 protein expression was located in theca cells around follicles, interstitial cells, and some luteal cells. Nesfatin-1 was also expressed in oocytes in some follicles, depending on follicle maturity (Fig. 3G, H). In the testis, nesfatin-1 protein expression was detected in Leydig cells and interstitial cells between seminiferous tubules (Fig. 3J, K).

DISCUSSION
Nesfatin-1 protein produced by neurons in the hypothalamus and secreted into cerebrospinal fluid controls appetite and energy metabolism (Oh-I et al., 2006). It was recently reported that nesfatin-1/NUCB2 mRNA is expressed in gastric organs such as the stomach and pancreas (Stengel et al., 2010; Zhang et al., 2010) and in adipose tissues of humans and rodents (García-Galiano et al., 2010). We showed that nesfatin-1/NUCB2 protein and mRNA are expressed in mouse reproductive organs. In addition, nesfatin-1 binding sites have been detected in mouse ovary and testis (Kim et al., 2010; Kim et al., 2011). Given these previous results, we hypothesized that nesfatin-1/NUCB2 are expressed in various organs in the body. In the current study, we demonstrated that nesfatin-1/NUCB2 protein and mRNA are expressed in various tissues of female and male mice. Furthermore, we analyzed the relative expression levels of nesfatin-1/NUCB2 in various organs of the body and compared the expression levels between female and male mice.
To examine mRNA expression of NUCB2 in various organs of female and male mice, we performed conventional PCR using total RNA extracted from the cerebrum, hypothalamus, pituitary, stomach, heart, spleen, lung, ovary, uterus, testis, epididymis, and muscle. This showed that NUCB2 mRNA was expressed in all organs analyzed. The relative level of NUCB2 mRNA expression in each organ could not be determined by conventional PCR. Therefore, we measured the level of NUCB2 mRNA expression in various organs by real-time PCR. We confirmed that the pattern of NUCB2 mRNA expression was similar in female and male mice. In both female and male mice, a large amount of NUCB2 mRNA was detected in the pituitary. In addition, NUCB2 mRNA was also highly expressed in the stomach, lung, and reproductive organs.
Next, we investigated the level of nesfatin-1 protein expression in various organs of female and male mice by western blotting. Nesfatin-1 protein was detected in all organs tested. These data corresponded with the real-time PCR analyses of NUCB2 mRNA expression. The level of nesfatin-1 protein was higher in the pituitary than in the hypothalamus in both female and male mice. Nesfatin-1 protein was much more abundant in reproductive organs than in the stomach and adipose tissue. Recent reports showed that nesfatin-1/NUCB2 protein and mRNA are expressed in the anterior pituitary (Goebel et al., 2009; Foo et al., 2008). These results raise the possibility that nesfatin-1 produced in the brain controls metabolic homeostasis not only via the hypothalamus, but also via the pituitary gland. In other words, nesfatin-1 protein produced by the pituitary may be transported to various organs in the body through the bloodstream and function as a hormone, similar to many other hormones produced by the pituitary. However, more studies are required to elucidate the role of nesfatin-1 in the pituitary gland.
Our data showing that nesfatin-1 protein and NUCB2 mRNA were expressed in mouse stomach are consistent with previous reports (Zhang et al., 2010). X/A-like cells in the gastric mucosal layer secrete ghrelin, a protein that stimulates appetite, and these cells also express nesfatin-1 (Stengel et al., 2010). Our data also showed that nesfatin-1 protein and NUCB2 mRNA were highly expressed in the fat of male and female mice. Nesfatin-1 protein was recently confirmed to be expressed in the fat of human and rodents, with expression higher in subcutaneous fat than in visceral fat (Ramanjaneya et al., 2010). The present study demonstrated that nesfatin-1/NUCB2 is expressed in reproductive organs. Several reports have demonstrated the localization of nesfatin-1 protein in interstitial cells, including Leydig cells, in rat and mouse testis (Gonzalez et al., 2010; Kim et al., 2010), and confirmed that nesfatin-1 protein is expressed in mouse ovary and uterus (Kim et al., 2010; Kim et al., 2011). These results suggest that nesfatin-1 plays an important role in reproductive organs involved in steroidogenesis.
Therefore, we next investigated which cells express nesfatin- 1 protein by performing immunohistochemical staining. In the ovary, nesfatin-1 protein was localized in theca cells around follicles, interstitial cells, and some luteal cells. Nesfatin-1 protein was expressed in some oocytes in mature follicles. The roles of nesfatin-1 in oocytes are unknown, although this localization of nesfatin-1 is assumed to be associated with the level of oocyte maturity or apoptosis. In the testis, nesfatin-1 was localized in interstitial cells and Leydig cells in seminiferous tubules. The function of nesfatin-1 in testicular cells is unclear; however, our immunehistochemical staining results raise the possibility that nesfatin-1 is involved in steroidogenesis to produce testosterone in Leydig cells (Ewing et al., 1983). In our previous study, we detected nesfatin-1 binding sites in the testis. Nesfatin- 1 protein binds to interstitial cells, including Leydig cells, and boundary cells in the tunica albuginea (Kim et al., 2010). The finding that Leydig cells express nesfatin-1 protein and harbor its binding sites indicates that nesfatin-1 plays an important role in testicular function as a local regulator, similar to autocrine signaling in the testis.
In this study, we reported for the first time that nesfatin- 1/NUCB2 is expressed in various tissues of male and female mice, and are highly expressed in the pituitary and reproductive organs such as the testis, ovary, uterus, and epididymis. These results suggest that nesfatin-1 plays an important role as a local regulator in various organs. Further studies are required to elucidate the mechanism of action of nesfatin-1 in each organ.