
INTRODUCTION
Actin is an abundant and highly conserved intracellular protein that exists either in a monomeric globular actin (G-actin) or as filamentous actin (F-actin) composed of G-actin monomers (Dominguez & Holmes 2011). F-actin, a major component of the actin cytoskeleton, assembles into bundles or branched networks and participates in various cellular processes, including cell motility, growth, polarity, and migration (Rajan et al., 2023). Despite its ubiquitous presence in cells, numerous studies have highlighted its localization in and around cilia, and the critical role of the actin cytoskeleton in ciliogenesis (Brücker et al., 2020). In primary cilia, mutations in ciliopathy genes that lead to increased actin stress fibers disrupt ciliogenesis, while treatment with actin polymerization inhibitor like cytochalasin D enhances ciliogenesis (Kim et al., 2010; Smith et al., 2020). Conversely, cytochalasin D treatment does not affect cilia morphology but disrupts cilia polarity and basal body spacing in multiciliated cells (MCCs), suggesting distinct roles for actin dynamics in primary cilia and MCCs.
The actin depolymerizing factor family, comprising destrin (Dstn) and coflins (Cfl1 and Cfl2), regulates actin dynamics by severing filaments and promoting depolymerization (Bamburg 1999; Bamburg & Wiggan 2002). Several studies have implicated Cfl1 in ciliogenesis; its depletion induces ciliogenesis in mammalian cells, and Cfl1 knockdown leads to ciliary defects in zebrafish (Kim et al., 2015; Zhang et al., 2016). Additionally, Cfl1 cooperates with Vangl2 to initiate planar cell polarity in mouse node cilia (Mahaffey et al., 2013). However, the role of Dstn in ciliogenesis remains unexplored. Understanding the regulatory mechanisms of ciliogenesis vi actin dynamics regulators is crucial for better insight into cilia biology in both monociliated cells and MCCs.
Therefore, this study aimed to elucidate the role of Dstn in ciliogenesis using Xenopus laevis and human retinal pigment epithelial cells (hRPE1). We performed loss-of-function experiments using morpholino oligonucleotides (MOs) in developing Xenopus embryos and siRNA in hRPE1 cells. Our results show that inhibition of dstn in Xenopus led to an increase in the number of MCCs and significantly disrupted the polarity of basal bodies in MCCs. In contrast, dstn knockdown in both hRPE1 cells and the Xenopus’ neural tube inhibited primary cilia elongation without influencing basal body docking. These findings suggest that Dstn regulates ciliogenesis by modulating actin dynamics in both primary and MCCs.
MATERIALS AND METHODS
Experiments were conducted strictly following the guidelines of the Animal Care and Use Committee, consistent with international laws and policies (National Institute of Health) Guide for the Care and Use of Laboratory Animals, publication no. 85-23, 1985. The Institutional Review Board of Kyungpook National University in Korea approved the experimental use of amphibians (2021-0017). All members of the research group received training for the appropriate care and use of experimental organisms.
X. laevis eggs were obtained and in vitro fertilized using standard methods as previously described (Sive et al., 2007). For mRNA injection or in situ hybridization using RNA probe, dstn and alpha-tubulin primers were designed based on National Center for Biotechnology Information (NCBI) and Xenbase sequences. Plasmids were constructed for mRNA synthesis using the pCS107 vector with BamH1 and Xho1 restriction enzymes. The RNA probe sequence was labeled with digoxigenin (DIG, Roche, Basel, Switzerland) and inserted into a T-easy vector (pGEM, Promega, Madison, WI, USA). The pCS107 and T-easy vectors were linearized with the Apa1 restriction enzyme, and capped mRNAs were synthesized using the SP6 mMessage mMachine kit (Invitrogen, Carlsbad, CA, USA). DIG-labeled RNA probes were generated using the T7 mMessage mMachine kit (Invitrogen).
dstn MO with the nucleotide sequence (5′-TCCGAACACCTGATGCCATTGTTGA-3′) was purchased from Gene Tools. For the rescue experiments, mutant constructs (dstn*) not recognized by dstn MO were sub-cloned into the pCS107 vector. Embryos at the two-cell stage were microinjected with mRNAs (dstn, dstn*, centrin-RFP, clamp-GFP) and dstn MO.
Embryos at stage 32 were fixed in MEMFA (4% paraformaldehyde, 0.1 M MOPS (pH 7.4), 1 mM MgSO4, and 2 mM EGTA) overnight at 4°C. Antisense digoxigenin (DIG)-labeled probes were generated from plasmids containing dstn or alpha-tubulin linearized by ApaI. The probes were synthesized using the mMESSAGE mMACHINE™ SP6 Transcription Kit (Invitrogen) and detected using an alkaline phosphatase-labeled anti-digoxigenin antibody (1:1,000, Roche) and NBT/BCIP staining solution (Roche).
Total RNA was extracted from hRPE1 cells, and cDNAs were synthesized using the PrimeScript™1st strand cDNA synthesis kit (Takara, Shiga, Japan). RT-PCRs were performed using Emerald Amp PCR Master Mix (Takara) and specific primers listed in Table 1.
| Gene | Forward primer | Reverse primer |
|---|---|---|
| GAPDH | 5′-CCACTCCTCCACCTTTGACG-3′ | 5′-CCACCACCCTGTTGCTGTAG-3′ |
| Human DSTN | 5′-GGCACCAGAACTAGCACCTC-3′ | 5′-AGCCCGATTGAGATCTTCTGG-3′ |
Embryos were anesthetized using 0.1X benzocaine, fluorescent microbeads were added to the culture media, and videos were recorded. The velocity of fluorescent beads was measured using the fluorescent bead measurement program, Tracker 6.0.
Human telomerase-immortalized retinal pigmented epithelial cells (hRPE1) were cultured at 37 °C and 5% CO2 in DMEM/F12 media (Thermo Fisher Scientific, Waltham, MA, USA) containing 10% fetal bovine serum (Welgene, Gyeongsan, Korea) and 100 U/mL streptomycin/penicillin. dstn siRNA (Table 2) and control siRNA (Bioneer, Daejeon, Korea) were transfected into hRPE1 cells for 24 hrs using Lipofectamine RNA iMAX (Invitrogen). Next, the culture medium was replaced with a serum-free medium for cells with 90% confluency and cultured for an additional 48 hrs. Finally, DSTN-flag plasmids were transfected using Lipofectamine 2000 (Invitrogen).
| Target gene | Sequence 5’-3’ |
|---|---|
| DSTN siRNA #1 | CAGAACUAGCACCUCUGAA |
| UUCAGAGGUGCUAGUUCUG | |
| DSTN siRNA #2 | CUCCUUCAGAAGUUUGCUU |
| AAGCAAACUUCUGAAGGAG |
For immunofluorescence, Xenopus embryos were fixed with MEMFA, and hRPE1 cells were fixed in 4% paraformaldehyde or methanol, depending on antibodies. Primary antibodies used were anti-acetylated tubulin (1:1,000, Sigma-Aldrich, St. Louis, MO, USA), CEP164 (1:1,000, Proteintech, Rosemont, IL, USA), IFT20 (1:200, Proteintech), and Rab11 (1:500, Invitrogen). The fluorescent-labeled secondary antibody was Alexa Fluor (1:2,000, Invitrogen). For F-actin staining, Xenopus embryos at stage 32 were fixed in MEMFA and stained with Alexa Fluor 488 phalloidin (1:1,000, Invitrogen). Images were obtained using an Olympus FV1200 confocal microscope.
Cell lysates were prepared using RIPA lysis buffer (150 mM NaCl, 10 mM Na2HPO4, 0.5% sodium deoxycholate, 1% NP40) supplemented with 1 mM PMSF (phenylmethylsulfonyl fluoride), 5 mM sodium vanadate, and 1 mM protease inhibitor cocktail (Roche). Lysates were loaded onto 12% SDS-polyacrylamide gels and transferred to a nitrocellulose membrane. The membrane was blocked in a 5% skim milk TBST buffer for 1 h at room temperature. Proteins were detected using polyclonal anti-DSTN (1:1,000, https://www.antibodies-online.com), and GADPH (1:1,000). The secondary antibody used was anti-rabbit IgG HRP-linked antibodies (1:2,000, Santa Cruz, Dallas, TX, USA).
ImageJ software (National Institutes of Health) was used for the analysis of confocal imaging and optical microscope imaging data. Statistical analyses were conducted by GraphPad Prism version 9. The results are presented as the means±SE from the indicated number of independent experiments in graphs (n). The data were analyzed using the unpaired t-test and the level of significance was considered as * p<0.05.
RESULTS
To investigate the role of Dstn in MCC formation during Xenopus embryogenesis, we conducted loss-of-function studies using dstn MO. We observed MCCs across larval skin through whole¬mount in situ hybridization (WISH) analysis, using an alpha-tubulin probe. The analysis revealed a significant increase in the number of MCCs in dstn knockdown embryos compared to control embryos (Fig. 1A). This increase in MCCs was effectively rescued in embryos injected with dstn* mRNAs (Fig. 1B). In addition to ciliogenesis, we also investigated whether dstn knockdown affects the function of multicilia, which are evolutionarily conserved structures that beat synchronously and coordinately to direct fluid flow across vertebrate epithelium (Brooks & Wallingford, 2014; Walentek et al., 2014) (Fig. 1C). Fluorescent microbeads were used to the velocity of cilia-driven fluid flow, using fluid flow tracker (Kim et al., 2023). The velocity of microbeads was significantly reduced in morphants compared to control embryos (Fig. 1D; Supplementary videos 1 & 2). These results suggest that Dstn plays a crucial role in MCC development and the synchronized beating of cilia during Xenopus embryogenesis.
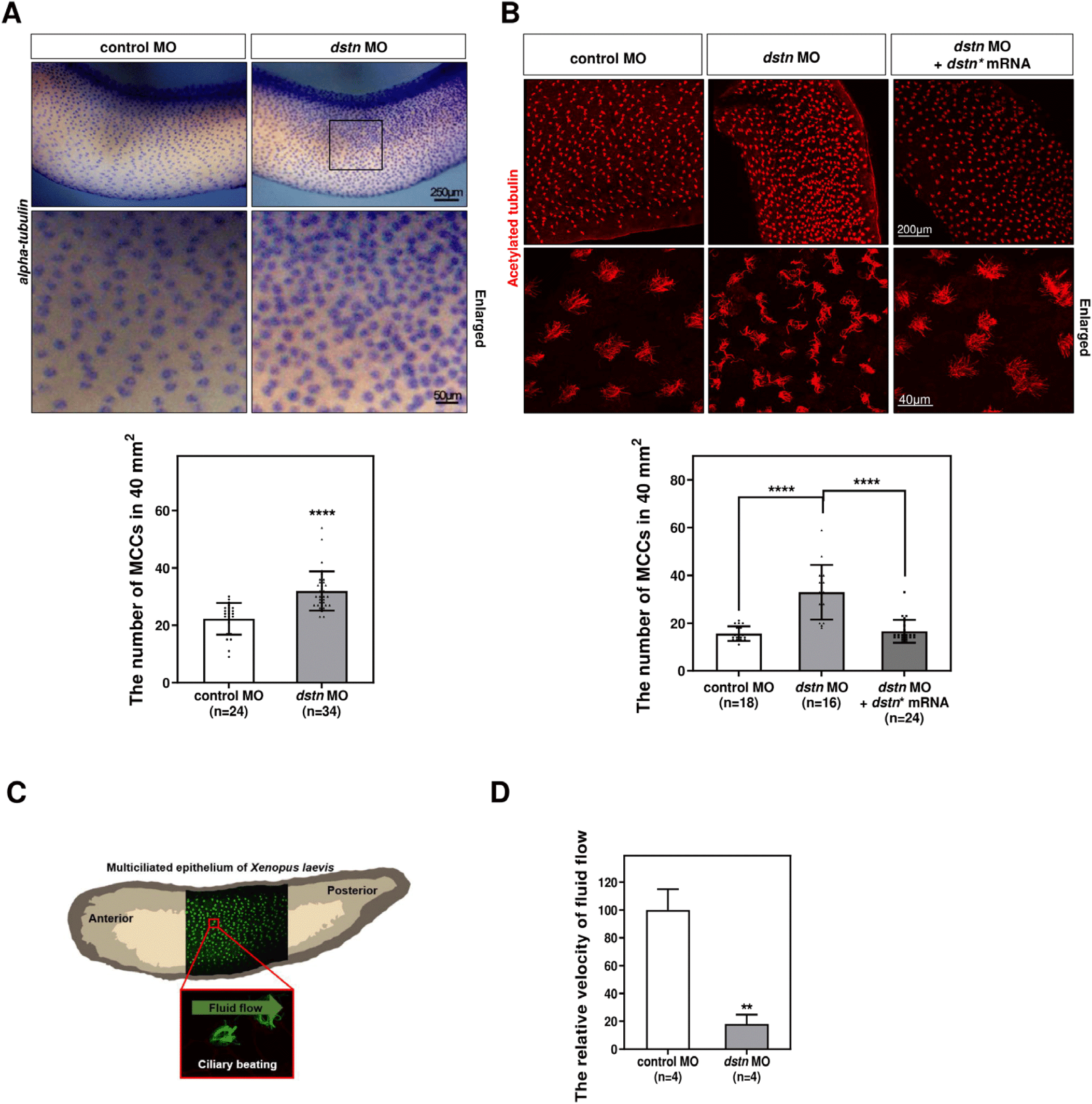
Given the close relationship between microtubule-based cilia and actin filaments, we hypothesized that both cytoskeletons are intrinsically linked during ciliogenesis (Werner et al., 2011). MCCs require an apical actin network for the docking of basal bodies at the apical cell surface during ciliogenesis (Smith et al., 2020). Compelling evidence supports the involvement of DSTN in actin filaments depolymerization (Hotulainen et al., 2005; Kumar et al., 2012; Zhang et al., 2016). To assess whether Dstn affects actin dynamics during MCC development, we visualized actin filaments on the larval epithelial surface using phalloidin staining. The phalloidin intensity (green) in MCCs labeled with acetylated tubulin (red) was significantly higher in dstn morphant embryos compared to control embryos (Fig. 2A). Additionally, confocal microscopic images revealed that actin fibers accumulated on the apical surface of dstn morphants along the z-axis (Fig. 2A).
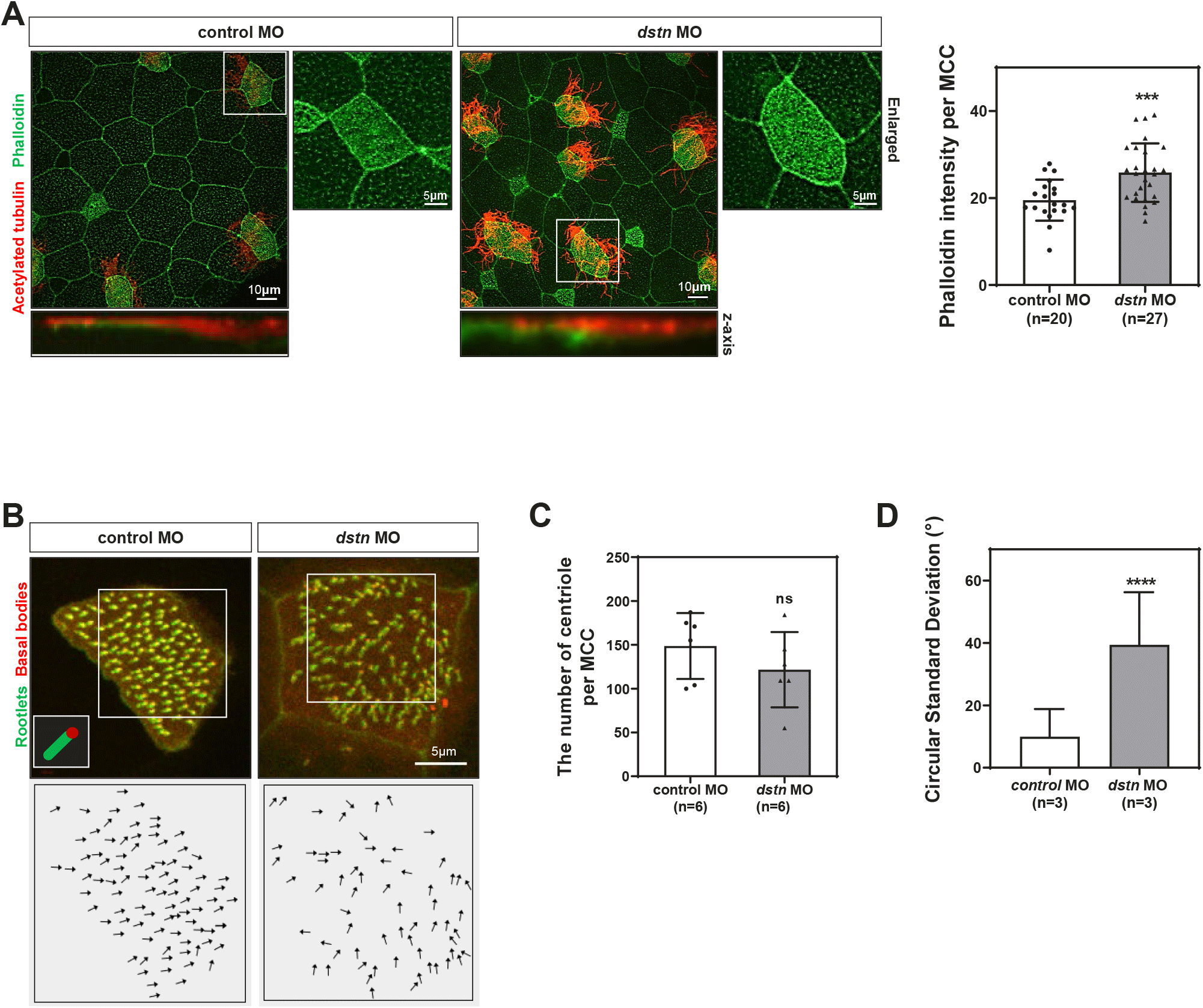
The docking of basal bodies at the cell membrane and the establishment of polarity are necessary for coordinated ciliary beating to generate fluid flow (Boisvieux-Ulrich et al., 1985; Mitchell et al., 2007). Basal body docking in MCCs requires an apical actin network (Sedzinski et al., 2017). Additionally, actin and microtubules regulate different aspects of planar cell polarity in MCCs by influencing ciliary beating (Werner et al., 2011). To better understand the roles of Dstn in basal body docking and polarity, we used fluorescent-tagged Centrin (red) and Clamp (green) to visualize basal bodies and rootlets respectively. Dstn knockdown led to basal body clumping and a loss of ciliary polarity (Fig. 2C and D). Quantification of basal body orientation, based on the basal body/rootlet angle relative to the body axis, showed a significant disruption in directionality following dstn knockdown (Fig. 2D). These results suggest that inhibition of Dstn causes an accumulation of F-actin beneath cilia in MCCs, disrupting basal body polarity and, consequently, the overall process of ciliogenesis.
There is ample evidence supporting the role of actin destabilization in primary ciliogenesis, while MCC formation requires the apical actin network (Hotulainen et al., 2005; Werner et al., 2011). However, the roles of actin-depolymerizing in the regulation of primary and multicilia formation remain controversial, warranting further investigation (Zhang et al., 2016). To clarify the role of Dstn in multicilia and primary cilia formation, we studied Xenopus embryos and human retinal pigmented epithelial (hRPE1) cells.
In the developing Xenopus neural tube, cells possess primary cilia that mediate signaling necessary for neural tube patterning (Walentek & Quigley, 2017). To test whether Dstn affects neural tube primary cilia, we used immunofluorescence on head sections. Immunostaining with acetylated tubulin revealed that the primary cilia of dstn-depleted embryos were significantly shorter than those of control embryos (Fig. 3A). To further assess the effects of DSTN inhibition on primary cilia, we transfected hRPE1 cells with DSTN siRNA and induced primary cilia formation through serum starvation. RT-PCR and western blot analyses confirmed the efficiency of DSTN siRNA, showing substantial reductions in RNA and protein levels (Fig. 3B and C). Similar to Xenopus, DSTN depletion in hRPE1 cells led to a significant reduction in primary cilia length (Fig. 3D). Altogether, these findings demonstrate that the Dstn regulates primary cilia formation in the same context in Xenopus and hRPE1 cells.
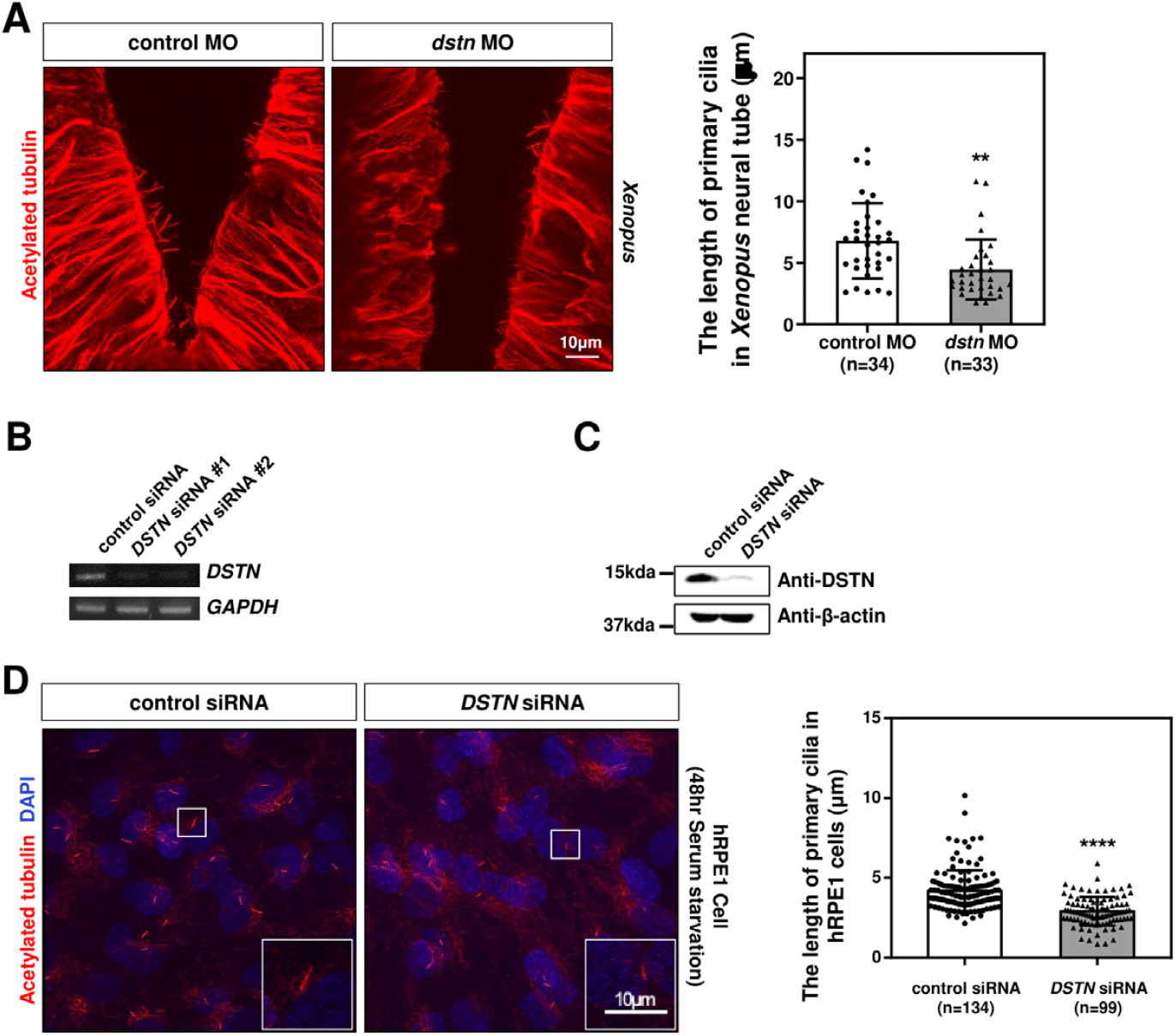
Ciliogenesis is a multistep process that includes the docking of ciliary vesicles, which is regulated by protein components such as Cep164 (distal centriolar protein) (Cao et al., 2012), Rab11 (ciliary vesicle trafficking protein) (Knödler et al., 2010), and IFT 20 (intraflagellar transport protein/ciliary membrane transport protein) (Zhu et al., 2017), followed by ciliary axoneme elongation (Nigg & Raff, 2009). To evaluate the role of Dstn in ciliary assembly, we transfected hRPE1 cells with DSTN siRNA, serum-starved the cells for 48 hrs, and performed immunostaining for Cep164, Rab11, and IFT20. Confocal images showed that DSTN depletion did not affect the localization or distribution of these basal body-associated proteins, but axoneme length was significantly reduced (Fig. 4A).
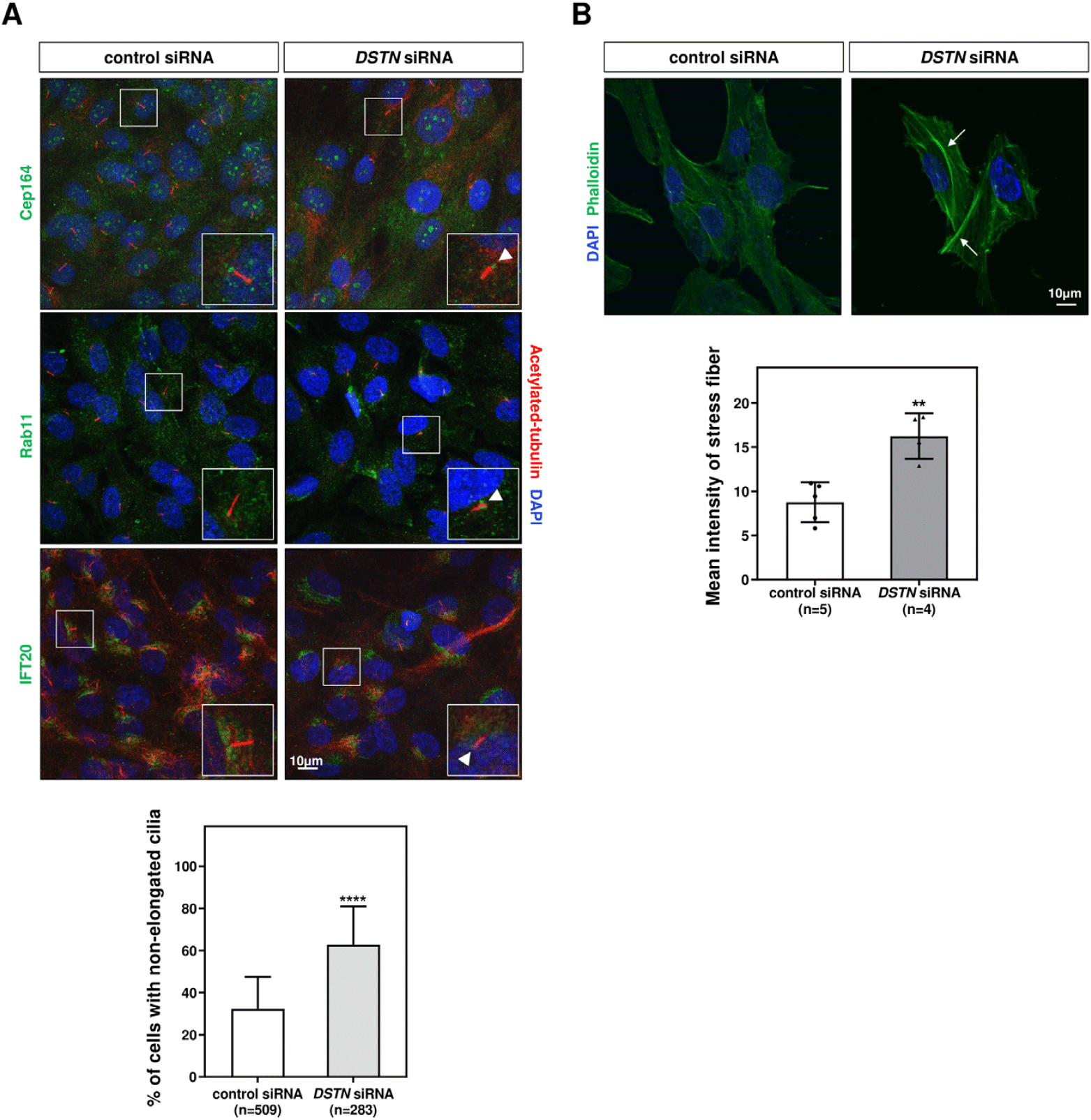
In earlier experiments, we demonstrated that Dstn influences actin filament accumulation during motile ciliogenesis (Fig. 2A). To determine whether Dstn similarly regulates actin dynamics in primary cilia, we performed immunofluorescence staining using phalloidin in DSTN-depleted hRPE1 cells. DSTN depletion resulted in thicker stress fibers compared to control siRNA-transfected cells (Fig. 4B). Given prior reports that increased stress fiber formation hinders primary cilia development, our results underscore the crucial role of Dstn in regulating primary ciliogenesis in hRPE1 cells.
DISCUSSION
Remodeling of the actin filament network provides insights into the regulation of ciliogenesis and cilium length (Pan et al., 2007). While CFL1, a member of the actin depolymerizing factors family, has been studied for its roles in ciliogenesis through the regulation of actin dynamics, the function of Dstn in ciliogenesis remains less understood. In this study, we used X. laevis as an in vivo model (Hayes et al., 2007) to demonstrate that Dstn governs cilia development and the polarized beating of cilia. Firstly, Dstn depletion increased the number of MCCs (Fig. 1A and B). Additionally, the loss of dstn resulted in distorted ciliary beating and a significant reduction in cilia-driven fluid flow (Fig. 1D). This indicates that Dstn is essential for organizing actin filaments in MCCs, as actin dynamics and ciliogenesis are intrinsically linked (Smith et al., 2020). As shown in Fig. 2A, the loss of dstn resulted in the accumulation of actin filaments at the apical surface of MCCs. However, this loss did not affect the docking and number of basal bodies but instead led to basal body clumping, significantly impacting their directionality and polarity (Fig. 2B–D). Our data suggest that Dstn governs the ciliogenesis of MCCs by affecting basal body polarity while not influencing basal body docking.
Further examination of Dstn’s role in primary cilia formation revealed that DSTN depletion leads to a reduction in primary cilia length in both Xenopus and hRPE1 cells (Fig. 3A and D). The shorter primary cilia after depletion of Dstn align with previous studies that highlighted the positive roles of CFL1 in ciliogenesis through the influence of LIMK2 and TESK1 modulation of YAP/TAZ activity (Kim et al., 2015). However, contrasting findings have also been reported, where CFL1 depletion increased the length of primary cilia in hRPE1 cells, influenced by NudC (Sedzinski et al., 2017). Thus, our results suggest that the regulation of ciliogenesis by Dstn involves the orchestration of a complex network and it can have both positive and negative roles in ciliogenesis. Understanding how cells integrate various signals to regulate ciliogenesis, particularly in reconciling these seemingly contradictory findings about actin depolymerizing factors’ roles in primary cilia length regulation, will be a key challenge for future studies.
Our investigation into the mechanistic details of Dstn in primary cilia regulation showed that while Dstn controls axoneme elongation, it does not affect the localization and distribution of basal bodies, mirroring its effects in MCCs (Figs. 2 and 4). Additionally, Dstn influences primary cilia formation by regulating actin stress fibers (Fig. 4B). Altogether, our data suggest that Dstn regulates ciliogenesis in both primary and motile cilia by modulating actin dynamics.
The paradoxical behavior of Dstn in primary cilia and multicilia can be explained by the distinct impacts of actin destabilization: promoting ciliogenesis in primary cilia, while having opposite effects in multicilia (Pan et al., 2007; Smith et al., 2020). In primary cilia, destabilized F-actin facilitates vesicle trafficking, which promotes ciliogenesis and increases ciliary length (Kim et al., 2010, 2015). Therefore, our findings that Dstn depletion reduces primary cilia length are consistent with previous reports (Wheway et al., 2015; Yeyati et al., 2017), indicating that Dstn positively regulates ciliogenesis by actin destabilization in primary cilia. The contradictory response observed in multicilia is again in agreement with reported data (Sedzinski et al., 2017) as multicilia formation requires an apical actin network for basal body docking and polarity establishment to allow synchronous ciliary beating (Mitchell et al., 2007; Pan et al., 2007; Kulkarni et al., 2018). F-actin stabilization and apical actin network support ciliogenesis by facilitating basal body docking in mouse tracheal epithelial cells, as the actin web expands the docking surface and spaces out the basal bodies across cell membrane (Werner et al., 2011; Sedzinski et al., 2017).
In conclusion, Dstn plays a significant role in ciliogenesis, regulating both primary and multicilia development. Our findings provide new insights into the roles of actin dynamics in ciliogenesis, contributing to a deeper understanding of this complex biological process.